|
Authors: Shuyana Deba, Javier Del Río and Sara Sanz Special collaboration: Álvaro Martínez Moro 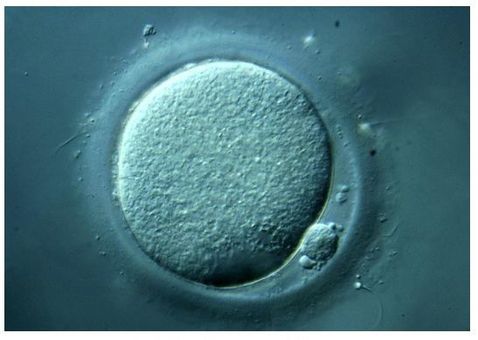 Figure 1. Metaphase II oocyte (1) Infertility affects millions of couples all around the world. In spite of the solutions to their problems reproductive technology can achieve, the efficacy is eventually limited by the number and the quality of the oocytes available from the woman. In actuality, such efficiency is determined by the ovarian reserve, the oocyte quality and the maternal age, among the most important factors (2). Diminished ovarian reserve (DOR) Since ovarian reserve defines the quantity and quality of the primordial follicle pool, diminished ovarian reserve (DOR) indicates a reduction in quantity in women of reproductive age. Consequently, it represents important cause of infertility in many couples. Moreover, DOR may be associated with low pregnancy rates and high pregnancy loss regardless of age, but further research is needed in order to fully understand its implications (3). Advanced maternal age It is well known that women’s fertility declines sharply after age 35 due to several factors, which include specific issues of reproductive organs (uterus and oviducts), general health and decreasing number and quality of oocytes over time. The oocyte pool starts to decline during foetal life and continues within the reproductive life of women. Oocyte quality also decreases as a consequence of the increased rate of aneuploidies observed with age: 74% at the age of 41–42, and up to 93% after the age of 42 (5). Advanced age is too associated with a reduction in the quality of the oocyte cytoplasm (ooplasm), which directly affects oocyte maturation (3). What are the main reasons for this reduction in ooplasm quality? Mitochondria are one of the most important organelles, which are affected in different ways (6,7): - Morphological and functional abnormalities - Mitochondrial swelling - Alterations in mitochondria's cristae - Vacuolization - Alterations of the membrane potential - Alterations of the metabolic pathways in cummulus cells, which may result in impaired mitochondria biogenesis during oogenesis. These effects are due to the higher ratio of mutation consequence of the proximity of these organelles to the respiratory chain, the inefficient repair mechanism and the exposure of histories. How these changes affect oocyte quality (8)? First of all, negative effects on chromosome segregation have been observed as a result of a decreasing ATP concentration (9,10). Additionally, defects have been found in different signalling pathways such as Ca2+ signalling, which affects fertilization and the subsequent embryo development (11). 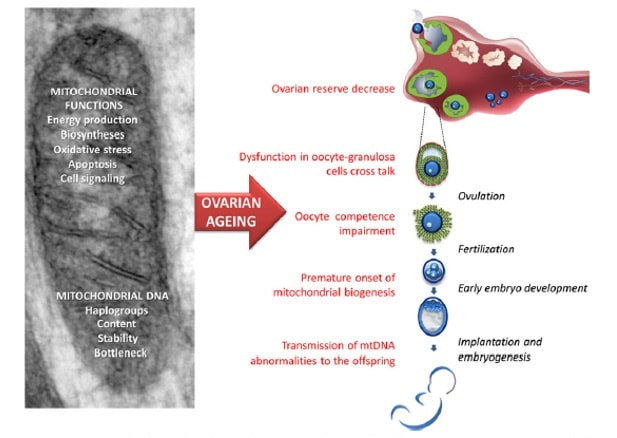 Figure 2. Overview of the multiple implications of mitochondria in ovarian ageing and their deleterious consequences on reproductive function (12) Nevertheless, different mitochondrial haplogroups should be taken into consideration. These have different bioenergetic functions, including production of reactive oxygen species (ROS) and mitochondrial coupling efficiency, aspects that might affect the oocyte longevity (13). Consequently, new techniques are being developed in order to increase the reproductive options in women with oocyte problems. Recently, one of these techniques that have been highly treated in the media is the development of additional viable oocytes from polar body genomes (2). HOW DOES TRANSFER OF POLAR BODY GENOME WORK? Originally, the transfer of polar body has been applied to cases of infertility with a genetic cause, such as the presence of mitochondrial diseases. These cases can be treated with the use of donor oocytes in clinical practice. Additionally, another application is the formation of human metaphase II (MII) oocytes, which increases the number of available oocytes for an assisted reproduction cycle (2). Two specific combined steps are needed. First, the donor oocyte spindle is removed, which requires the utilization of polarized light. Once located, it will be biopsied, obtaining an enucleated oocyte (14,15,16). Secondly, the patient polar body is biopsied, provided elimination of the spindle apparatus has been confirmed. Once both processes have been performed, the last step is the introduction of the polar body genome inside the enucleated oocyte (17). 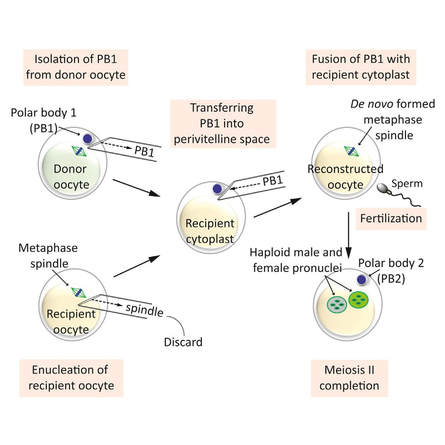 Figure 3. Polar body genome transfer technique (2) FUNCTIONAL HUMAN OOCYTES GENERATED BY TRANSFER OF POLAR BODY GENOMES Hong Ma and his group have tried to test the efficiency and possible limitations of this technique (ref). The main objective to be achieved was the formation of spindles resembling those typical of MII oocytes, including the appropriate chromosome dosage. HOW EFFICIENT IS THIS TECHNIQUE? Although DAPI staining demonstrated that all polar body nuclear transfer (PBNT)-oocytes contained spindle-chromosome complexes, only two of five experimental oocytes formed metaphase spindles similar to intact MII oocytes. This low number may be due to residual meiotic activity in enucleated human MII oocytes, which is sometimes not enough to induce formation of normal MII-like spindles. For a different cohort of oocytes, the rate of successful fertilization was 76%, still slightly lower than control oocytes. Furthermore, 42% of embryos reached blastocyst stage, indicating that most of the PBNT-oocytes were capable of completing the second meiotic division. Short tandem repeat (STR) analysis revealed that two sampled PBNT-blastocysts contained normal diploid chromosomes, determining that these embryos were completely viable. WHAT CAN BE CONCLUDED? • Polar body genome transfer seems to be a significant technique for the improvement of assisted reproductive technology (ART) outcomes and pregnancy rates, particularly for women with decreased ovarian reserve and low response to stimulation. • The cytoplasm from young donor oocytes may reduce incidences of low cytoplasmic oocyte quality. • It could provide an additional technique to support mitochondrial replacement therapy. Nevertheless, this technique is not suitable for women who cannot produce mature oocytes, typical profile of ART patients. Additionally, incidences of aneuploidy resulting from errors in mitosis or in the second meiotic division may still occur because of women advanced age. Larger datasets from this technique are needed to confirm its efficacy and safety. Also, improving preimplantation genetic screening (PGS) is critical before eventual clinical application. REFERENCES:
4 Comments
Authors: Shuyana Deba, Javier Del Río and Sara Sanz 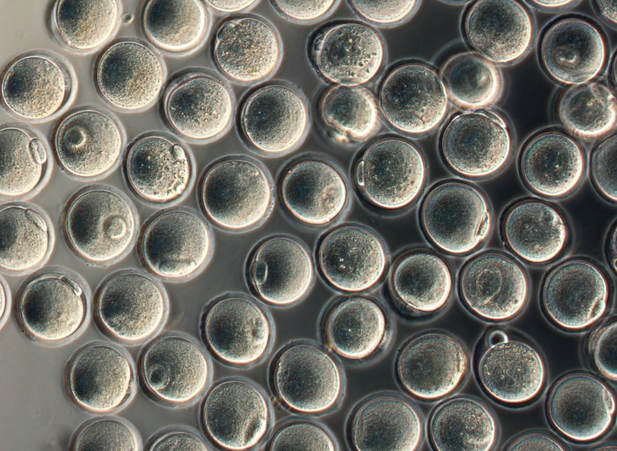 Figure 1: MII oocytes obtained in vitro (1) Some weeks ago, Dr. Hayashi's group published a study in Nature in which they described how to generate functional mouse eggs from cultured stem cells. To better understand this experiment, it is necessary to explain the natural ovarian cycle. Although the experiment was carried out in mice, we believe these experiments could be of great importance in humans in the near future. Therefore, the following description applies to human ovarian cycle. GONADAL DEVELOPMENT IN THE EMBRYO As the yolk sac begins to incorporate into the embryo, germ cells migrate along the dorsal mesentery of the hindgut to the gonadal ridges, which they reach by the end of the fourth or early fifth week of development (2). Around week 7 of gestation, gonadal ridges undergo proliferation to form primitive sex cords, to which primordial germ cells (PGCs) are associated (Fig. 2). In the absence of both expression of the SRY gene product and male sex hormones, differentiation into female organs is determined (3,4). Whereas the origin of the follicular cells of the ovary remains unclear, likely candidates are cells from the coelomic epithelium and the mesonephros. The follicular cells eventually associate with the PGCs to form primordial ovarian follicles (3,4). ORIGIN OF GERM CELLS Human oocytes are derived from round PGCs that can be identified in the wall of the yolk sac as early as 24 days after fertilization. Once the gonad develops into an ovary, PGCs become oogonia, and mitotic divisions go on up to the seventh foetal month, and cease at some point shortly before birth (5). By the 8-9th week after fertilization, some oogonia enter into prophase I of meiosis, thus becoming primary oocytes. Meiosis starts in the deepest part of ovary (medulla) and progresses towards the cortex. Some time after it has been incorporated into a primordial follicle, the oocyte arrests after diplotene, at a particular stage called dictyate or dictyotene. This means the oocyte enters into a long quiescence period that begins before birth and ends up either resuming after the LH surge (leading to ovulation) or in follicular atresia (3). Even though the experiment by Hayashi's group has just been successfully performed in mice, the purpose of the investigation is eventually applying it to humans. 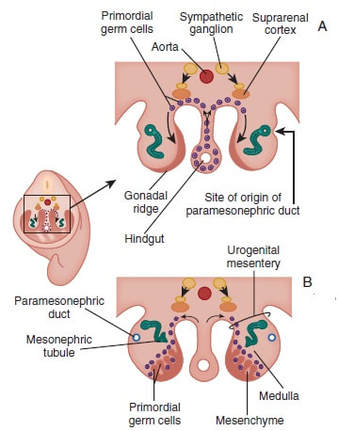 Figure 2: Gonadal development. C: Transverse sections showing the gonadal ridges and the migration of primordial germ cells into the developing gonads; D: transverse section through a 6-week-old embryo, showing the primary sex cords and the developing paramesonephric ducts (3) FOLLICULOGENESIS AFTER BIRTH Once placental oestrogen disappears, gonadotrophin levels begin to increase after birth. This increment will last 12-24 months postpartum in women. Thus, the ovaries remain inactive until the beginning of pubertal stage, moment when they will start showing response to GnRH pulsatility (6). By this time, ovaries contains germ cells at the primordial follicle phase, which are composed of small immature oocytes arrested at dictyate, and surrounded by a single layer of flat squamous granulosa cells (7). WHAT HAPPENS AFTER PUBERTY? Once sexual maturity is reached, multiple primordial follicles are activated from their quiescent state, resuming folliculogenesis by cohorts every menstrual cycle. During follicle maturation, follicles are subjected to activin, AMH, FSH, inhibin, LH, TGFβ and a multitude of other molecules that control the process. At the beginning of this process, as the oocyte resumes meiosis follicular cells proliferate and surround the oocyte, which develops into a primary follicle (7,8,9). Briefly, transition into the secondary or antral follicle stage involves the alignment of the stroma around the basal lamina and the development of an independent blood supply. Follicular cells of the underlying stroma, the granulosa cells, subsequently differentiate into an inner and an outer theca layers, the latter supporting this blood supply. The production of follicular fluid by these cells promotes the formation of the antrum in pre-antral follicles (10). Progression through antral follicle development is promoted by FSH and coincides with (7): - Completion of oocyte growth. - Acquisition of competence to complete meiosis (dependent on fertilization). - Continued granulosa cell proliferation. - Differentiation of cummulus and mural granulosa cells upon formation of the follicular antrum. Response to FSH causes growth of the follicle by proliferation of follicular cells and coalescence of the astral cavities, thus generating a follicle whose volume keeps increasing. The final stage of folliculogenesis takes place when the Graafian follicle reaches its maximum size (around 20 mm diameter). The LH surge triggers the ovulation of a metaphase II-arrested oocyte (along with the cummulus), capable of being fertilized and able to support embryonic development (7). Although there are notable differences between species, in humans, usually only one follicle is capable of completing growth and maturation up to ovulation (preovulatory), due to its more efficient response to FSH in a "competitive manner" against the rest of developing follicles. The one follicle will progress until subsequent fertilization (dominant follicle), whereas the remaining follicles in the cohort will undergo atresia (8). 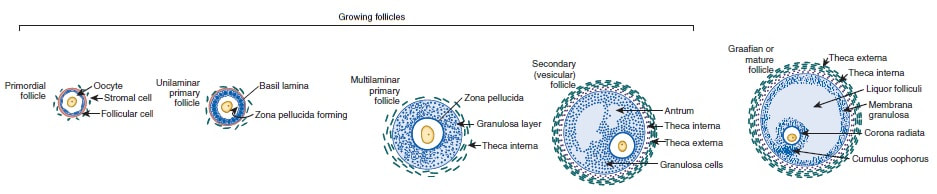 Figure 3. Shematic drawing of development of ovarian follicles (3) ABOUT PLURIPOTENT STEM CELLS For centuries, scientists have investigated how oocytes and sperm develop in mammals. Most of this work has been performed in non-human species, mainly the mouse. Human pluripotent stem cells (PSCs)-derived gamete research presents notable scientific value and potential, both for understanding basic mechanisms of gamete biology and for overcoming clinical problems (11). PSCs are currently used to answer important scientific questions such as the role of specific genes in early germ cell development, involved pathways, interaction between germ cells and supporting somatic cells, or partly or entirely in vitro PSCs-derived eggs and sperm (11). Some of the potential benefits of these new techniques could be (11): - Additional options for assisted human reproduction. - New ways to prevent and treat infertility, genetic diseases and some types of cancer. - Optimization of the number of oocytes obtained from women. "IN VITRO RECONSTITUTION OF THE ENTIRE CYCLE OF THE MOUSE FEMALE GERMLINE" (1) The authors have shown a method to reconstruct the entire process of mouse oogenesis in vitro, using embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) in order to obtain fully functional and mature oocytes. Why does this study mean so much? First of all, this is a big achievement in the field of regenerative medicine, through which it is possible to study the function of certain genes involved in oogenesis. Furthermore, it sets the ground for future similar studies and culture systems in other specie, and as such, it may result in a great advance in reproductive medicine. However, which are the limitations for this approach? The culture system requires somatic cells, which are obtained from embryos. So far, this is an ethical and legal issue in humans. A feasible solution would be to obtain the gonadal somatic cell-like cells from PSCs. The authors found that the differentially expressed genes (DEGs) were down-regulated in in vitro MII oocytes, whereas in vivo MII oocytes showed an increase in the number of transcripts. This maybe a consequence to the fact that, during in vitro growth (IVG) and in vitro maturation (IVM), oocyte development was jeopardized within a subset of (or perhaps all) the oocytes. Why? Misregulation of such genes, related to mitochondrial functions, might attenuate the potential of the in vitro MII oocytes. This problem could be solved by an analysis of the metabolic pathway, which could in turn provide information to refine both IVG and IVM. RESULTS, "PROBLEMS" AND "ACHIEVEMENTS" OF THE EXPERIMENTS (1)  As it can be seen concluded from above, it seems clear that further tests are still needed, which would help measure some aspects of the viability and functionality of this in vitro generated gametes.
REFERENCES 1. Hikabe O, Hamazaki N, Nagamatsu G, Obata Y, Hirao Y, Hamada N et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature. 2016;. 2. Baillet A, Mandon-Pepin B. Mammalian ovary differentiation–A focus on female meiosis. Molecular and Cellular Endocrinology. 2012;356(1-2):13-23. 3. Bolender DL, Kaplan S. Chapter 3 – Basic Embryology. In: Fetal and Neonatal Physiology. 2017. p. 23–39.e2. 4. Parker KL, Schimmer BP. Chapter 8 – Embryology and Genetics of the Mammalian Gonads and Ducts. In: Knobil and Neill’s Physiology of Reproduction. 2006. p. 313–36. 5. Racowsky C, Gelety TJ. Chapter 4 – The Biology of the ovary. Princ Med Biol. 1998;12:77–102. 6. David Vantman B, Margarita Vega B. Fisiología reproductiva y cambios evolutivos con la edad de la mujer. Revista Médica Clínica Las Condes. 2010;21(3):348-362. 7. Hutt K, Albertini D. An oocentric view of folliculogenesis and embryogenesis. Reproductive BioMedicine Online. 2007;14(6):758-764. 8. Green L, Shikanov A. In vitro culture methods of preantral follicles. Theriogenology. 2016;86(1):229-238. 9. Palermo R. Differential actions of FSH and LH during folliculogenesis. Reproductive BioMedicine Online. 2007;15(3):326-337. 10. Gougeon A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Annales d'Endocrinologie. 2010;71(3):132-143. 11. Mathews D, Donovan P, Harris J, Lovell-Badge R, Savulescu J, Faden R. Pluripotent Stem Cell-Derived Gametes: Truth and (Potential) Consequences. Cell Stem Cell. 2009;5(1):11-14. Authors: Javier Del Río and Sara Sanz  Figure 1. Illustration of a fetus (1). Genetic problems in the embryo are one of the most important causes of pregnancy loss and miscarriage. However, identifying embryo mosaicism as the cause of genetic problems during development is not an easy task. WHAT IS A MOSAIC EMBRYO? The term mosaicism refers to the presence of more than just one cell line, which present different chromosome count (1). Additionally, the most common situation in these cases is the presence of a mixture of distinct aneuploid cells, rather than of a variety between euploid and aneuploid cells. These are the embryos that may be at risk of misdiagnosis (2). There are four possible types of mosaic embryos (3,4): 1. Embryos with a mix of euploid and aneuploid cells in the trophectoderm (TE), and with aneuploid cells in the inner cell mass (ICM). 2. Embryos with a mix of euploid and aneuploid TE cells and euploid ICM. 3. Embryos with euploid TE cells and aneuploid ICM. 4. Embryos with aneuploid TE cells and euploid ICM. Even though there exists no specific cut-off to determine mosaicism, the Preimplantational Genetic Diagnosis International Society (PGDIS) suggests an embryo with more than 20% of aneuploid cells to be considered as mosaic. This means lower levels of mosaicism should be treated as normal (euploid) (5). It has been traditionally thought that only genetic problems in the oocyte or the sperm could be responsible for embryo mosaicism. Nevertheless, it is currently postulated that this also occurs during the first mitotic divisions, when maternal transcripts control the cell cycle of the early embryo (6). NORMAL INDICATIONS FOR CHROMOSOMAL TESTING TO DETECT EMBRYO MOSAICISM 1. Advanced maternal age. This is the most common cause for aneuploid problems (7). A recent study has shown that women between the ages of 35 and 43 years have more probabilities (an increase ranging from 28 to 78%) of presenting mis-segregation for the most clinically relevant aneuploidies, namely chromosomes 13,16,18,21 and 22 (8). 2. Severe male factor infertility. Even though levels of sperm aneuploidy are associated with increased levels of chromosomal abnormalities in embryos (9), such abnormalities could also arise from certain males who do not present any chromosomal abnormality a priori. Such could be specific cases of oligoastenozoospermic patients (10). 3. Recurrent implantation failure (RIF). In spite of the lack of specifications for such diagnosis, it is usually defined as the occurrence of three or more failed IVF attempts due to an unidentified cause. RIF is the usual diagnosis in those cases in which after a cumulative transfer of more than 10 good-quality embryos, the eventual result is IVF failure (7,11,12,13). 4. Recurrent miscarriage. The definition of this concept may vary for every country. However, generally speaking it can be defined as the occurrence of 3 or more consecutive miscarriages once pregnancy has reached at least 14 weeks (14). The main cause for this problem seems to be aneuploidy, which has been identified as the leading cause in a high percentage of miscarriages (15,16). 5. Previous trisomic pregnancy. Cases in which there has been a previous trisomic pregnancy entail higher probability of suffering from another aneuploid conception. Therefore, it is in this group of patients in which it would be beneficial to conduct a study to find out possible related causes (17). BIOPSY TECHNIQUES TO STUDY A CASE OF MOSAICISM Although there are different biopsy strategies, depending on the embryo stage, blastocyst biopsy is recommended over both polar body and blastomere biopsies in those cases in which mosaicism is suspected. Blastocyst biopsy is less invasive, it is possible to extract a higher number of cells, which increases the probability to confirm mosaicism, and it is cheaper than the other techniques due to the lower number of embryos required for biopsy (7). Furthermore, if cells with different chromosome complements are widely distributed throughout the trophectoderm, there might be a good chance of capturing a representative sample. If cells are clustered, mosaicism could not be easily detected, thus providing false normal results (18). 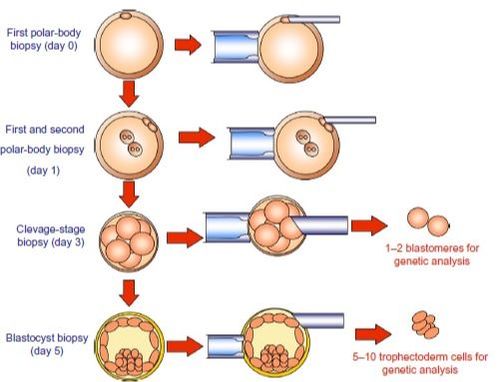 Figure 1. Different possible biopsy strategies (7) TECHNIQUES EMPLOYED TO DIAGNOSE MOSAICISM The earliest trials of PGD involved the use of karyotyping and PCR. By the mid-90s, the use of cytogenetic techniques such as FISH allowed for the progress of preimplantation diagnosis. This very own approach was later shown to impose important technical limitations to the analysis, and so it was encouraged the development of new technologies that could minimize the errors in diagnosis (19). Fluorescence in situ hybridization (FISH) It allows for the analysis and identification of chromosomes or chromosome fragments with 5-10 fluorescently labelled molecular probes from one cell (blastomere) (Fig. 2). This cell can be biopsied from day-3 embryos, it could belong to the trophectoderm from blastocysts or it could also be a polar body biopsied from an oocyte or a zygote. Therefore, PGS–FISH diagnosis is limited to the most common abnormalities involving chromosomes 13, 15–18, 21, 22, X and Y. Some studies of preimplantation embryos diagnosed by using this technique estimate a 5-7% error caused by mosaicism when embryos are reanalyzed. Additionally, FISH is being rapidly replaced by other DNA analysis methods with higher efficiency (20,21). 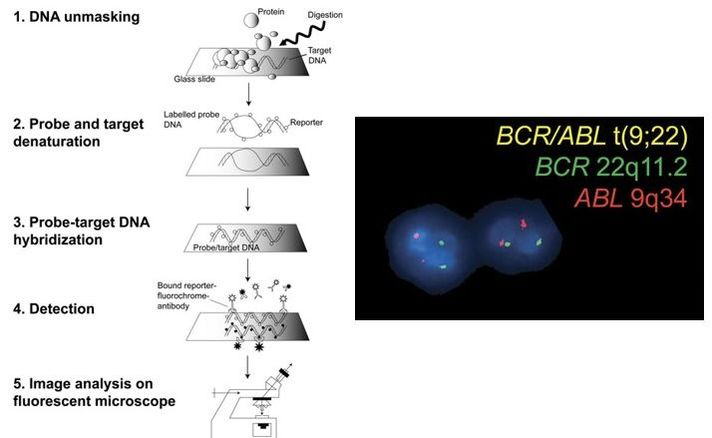 Figure 2. Fluorescence in situ hybridization (22) Array Comparative Genomic Hybridization (aCGH) This technique relies on whole genome amplification from one or more blastomeres (Fig. 3). It provides a quantitative analysis based on the comparison between the relative amount of tested DNA and the control DNA. Thus, chromosome imbalances such as aneuploidies, unbalanced translocations, deletions and duplications are easily detected. However, since balanced chromosome rearrangements such as reciprocal translocations or inversions do not affect copy number, such alterations cannot be identified (19,20). When blastomeres are analyzed by aCGH, the error rate measured is merely 2% (21,23). Notably, some studies have found that PGS-aCGH after blastocyst biopsy provides higher implantation and pregnancy rates than PGS-FISH (24). 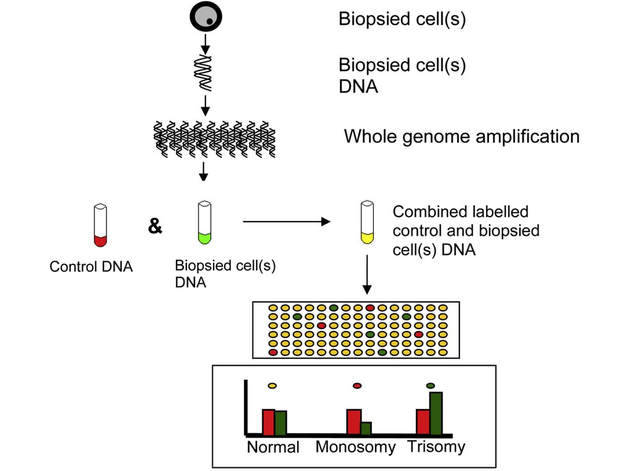 Figure 3. Array Comparative Genomic Hybridization (25) Next Generation Sequencing (NGS) Next Generation Sequencing belongs to the group of Massively Parallel Sequencing (MPS) methods that allow for parallel processing of an extremely large number of nucleic acid molecules (Fig. 4). As a result of sequencing on a microspace scale, it has been possible to drastically increase the amount of information collected during one test up to an entire human genome. Also, it is the only method that allows for analysis of all chromosomes (aneuploidies or translocations) and mutations responsible for any single-gene disease, just using one biopsy and in a single step (20). Although clinical results have documented high pregnancy rates following transfer of screened embryos, further data along with an extended use in clinical application are required to better define the role of NGS in PGS. Nevertheless, it seems that this method may actually lead to reduced costs per patient, thus allowing IVF couples a wider use of PGS for choosing the most competent embryo for transfer (26). 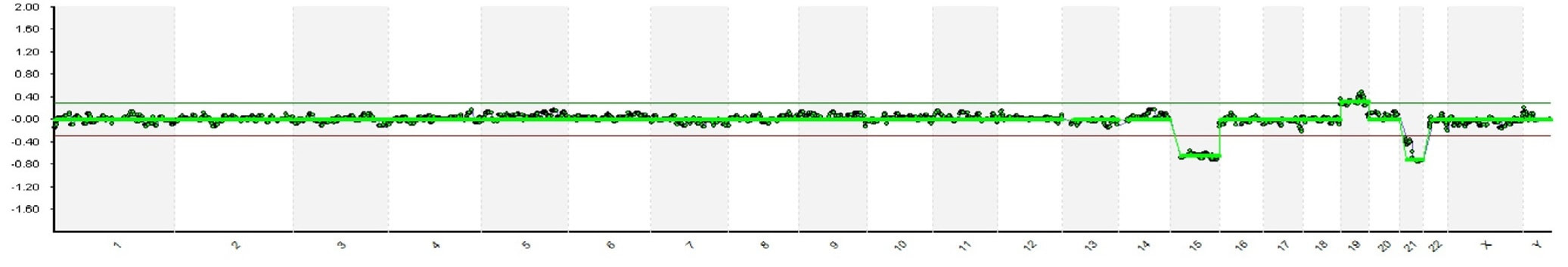 Figure 4. Next-Generation Sequencing (11) DIFFICULTIES WHEN MANAGING RESULTS The detection of mosaicism at an early stage does not mean that it will spread along embryo development (27). However, the utilization of chromosome identification techniques as part of the IVF process makes it possible to identify embryos “at risk of mosaicism’’ in order to select those that are suitable for transfer (18). Mosaic embryos are supposedly less competent than others due to a reduced implantation potential. Therefore, by discarding mosaic embryos implantation rates should be improved and, simultaneously, embryo loss rates reduced. Nevertheless, mosaic embryos may still have reproductive potential, and consequently they could still be viable. Furthermore, discarding embryos capable of producing healthy children will decrease pregnancy rates in those patients who get a low number of blastocysts in the pool of transferable embryos (18). It is important to take into account the reaction of the patients when they are informed about their embryos being at risk for mosaicism, what may entail genetic abnormalities, reduce implantation rates, increase loss risk and even diminish obstetrical and neonatal outcomes. However, there is not a simple answer when patients decide to transfer a mosaic embryo; either way, the obstetrical team should be informed for future screens (18). SUGGESTED GUIDELINES BY THE PREIMPLANTATINO GENETIC DIAGNOSIS INTERNATIONAL SOCIETY (PGDIS)
 Figure 5. Guidelines to priotitize mosaic embryos for transfer (5)
 Figure 6. Guidelines for the laboratory (5)
 Figure 7. Guidelines for the clinician (5) This article has been selected for publication in the Scientists in Reproductive Technologies (SIRT) Newsletter of The Fertility Society of Australia: DEL RÍO, J. and SANZ, S. (2017) Mosaic embryos are capable of producing healthy children. How to handle it? Fertility Society of Australia - SIRT Newsletter 4(20): 12-15.
REFERENCES
Authors: Javier Del Río and Sara Sanz  Although most of the genetic material of eukaryotic cells is located inside the nucleus, mitochondria are organelles that also possess a certain amount of DNA. Mutations in mitochondrial DNA (mtDNA) or nuclear genes involved in mitochondrial function are cause of infertility and diseases, not only in individuals, but also in their offspring. In these cases, one of the solutions known to be efficient in order to conceive and give birth to a healthy child is the "three-parent in vitro fertilization" approach. THE MITOCHONDRIAL GENOME IS MATERNALLY INHERITED During fertilization, mitochondria from sperm are normally eliminated by a ubiquitin-dependent mechanism. As a consequence, in case the father carries the mutation both his health and fertility could become affected, but never his offspring (2). By contrast, mitochondria in the oocyte must present a specific location and distribution pattern (which represents an actual sign of oocyte maturation), and they are solely inherited from the mother. WHY ARE MUTATIONS IN mtDNA OR IN NUCLEAR GENES INVOLVED IN MITOCHONDRIAL FUNCTIONS SO PROBLEMATIC? Mitochondria provide energy to the cells through oxidative phosphorylation, and so mutations in their genome mainly affect structures form the nervous system, heart, skeletal muscle, pancreas, gonads, colon, blood, kidney or liver (3). Why these structures? The higher the energy demand is, the higher the need for more mitochondria in the cells (4). Also, cells that present a slower division process are more likely to present some kind of mtDNA mutation (5). HOLOPLASMY vs. HETEROPLASMY The situation in which all cells from an individual contain identical mtDNA (mutated or otherwise) is known as holoplasmy. By contrast, heteroplasmy is defined as the condition in which part of the mitochondria from the same individual present a DNA content that is different from the other. These cases are the most common among patients affected by mitochondrial DNA diseases (6). WHY DOES HETEROPLASTY REPRESENT A PROBLEM? Even though heteroplasmy implies the presence of two different DNA contents in the cell, cells with great amounts of mutant (or affected by a specific condition) mitochondria respond to proliferate their entire DNA. This is why the percentage of mutant/affected mtDNA tend to increase in certain tissues (7). THE "BOTTLENECK EFFECT" AND THE "THRESHOLD EFFECT" During oogenesis, only a subset of molecules of mtDNA are eventually amplified and passed on to the offspring (8). This effect explains why it is possible to obtain homoplasmic individuals in just a few generations (2). Previous reports on human diseases caused by an mtDNA mutation have shown that the mutation needs to be present at a certain percentage in order to manifest pathological effects. Typically, this percentage should be higher than 60-80% (8,9), although it also depends on age, affected tissues, type of mutations, etc. (4) WHY TO APPLY THE "THREE-PARENT IN VITRO FERTILIZATION" APPROACH? It might be reasonable to think of other possibilities to treat patients suffering from mitochondrial diseases in order to achieve pregnancy. Rather than prenatal diagnosis or preimplantation genetic diagnosis, the "three-parent in vitro fertilization" technique because (4): In the first case: 1. It needs a uniform mtDNA distribution in the extra-embryonic and foetal tissues. 2. It needs the mutant DNA load to remain constant over time. 3. There must be a close relation between the severity of the disease and the amount of mutant DNA. As for preimplantation genetic diagnosis (6,9): 1. It is not applicable to patients with high levels of heteroplasmy. 2. It reduces but does not eliminate the risk of suffering from a mitochondrial disease-related condition. 3. The amount of tDNA found in blastomeres or the trophectoderm does not represent the whole embryo. 4. This approach is not an efficient diagnosis due to the combination of heteroplasmy and the "bottleneck effect". KNOWB TECHNIQUES TO BE USED FOR MITOCHONDRIAL REPLACEMENT Pronuclear transfer It involves the transfer of the two pronuclei from a zygote affected by diseased (or mutated) mitochondria into an enucleated zygote containing healthy mitochondria. Even though this technique has not yet been performed in humans, the efficiency of pronuclear transfer in mice has been adversely affected by descendants bearing high levels of carryover mtDNA (10). 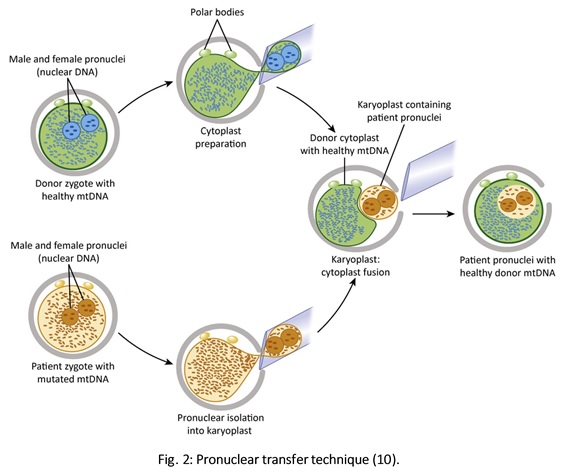 Polar body transfer Since the polar body has a lower proportion of mitochondria around it, this is currently considered the best method for preventing the transmission of mutated mtDNA on to the next generation (9,10). Embryos derived from polar body transfer support normal fertilization and are capable of producing live offspring in mouse. Polar body transfers leading to a minimal amount of affected mtDNA carryover have demonstrated the great potential of this technique for preventing inherited mitochondrial DNA diseases (9,10). When applied in mice, this technique has shown the best success rate so far due to the transfer of mitochondria being lowered to a minimum (9). 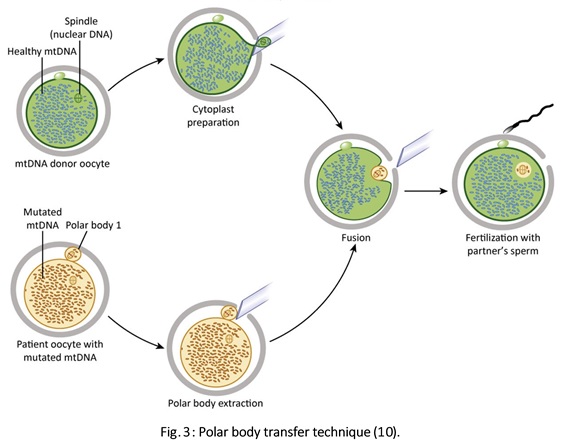 Spindle transfer This technique involves transferring the meiotic spindle along with the associated chromosomes, the spindle-chromosome complex (SCC), from an unfertilized oocyte with affected mitochondria into an enucleated healthy mitochondria-containing oocyte (11). This technique recently became popular when performed by Dr. John Zhang and his team, hitting the media within the latest weeks. However, potential problems could arise; just as for the previous techniques, the spindle is also surrounded by mitochondria, and so they could too be introduced into the ooplasm, thus causing heteroplasmy (12). 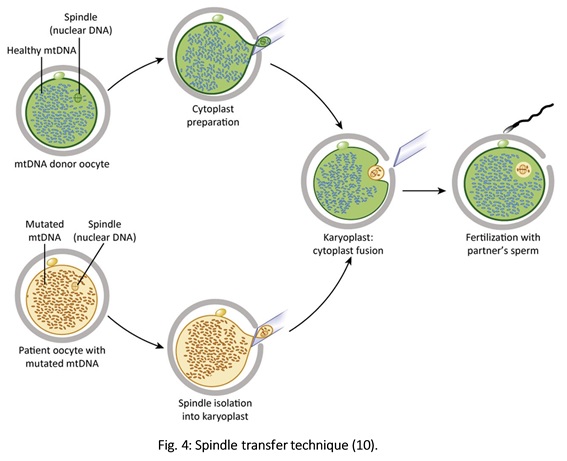 WHAT DOES LAW STATE REGARDING MITOCHONDRIAL REPLACEMENT? So far, scientific societies are very skeptical about experimental techniques. Thus, this particular approach is specifically prohibited by the Food and Drug Administration (FDA) in the US. It can only be performed in countries such as Mexico, where legislation is more flexible, or in UK, where it was approved for application in very specific cases (14). CURRENT DATA ON MITOCHONDRIAL REPLACEMENT As it has been previously mentioned, cases of cytoplasm transfer have been performed. In fact, there have been around 30-50 live births from this technique. However, newborns presented certain genetic defects, and so this technique was banned and replaced by pronuclear transfer and meiotic spindle transfer (14). Such data demonstrate the potential damage that could be inflicted to the embryo when performing these techniques (12,15). In addition to this, ethical issues must also be taken into account, which means the sole possibility of successfully applying a specific procedure does not imply its moral appropriateness. In order to guarantee so, a committee of experts should pronounce their opinions and reach a consensus about it. On a related note, long-term effects derived from these procedures are still unknown, and so it would be necessary to monitor all babies born through these techniques. This post has been published in the Scientists in Reproductive Technologies (SIRT) newsletter, a special interest group representing the scientific membership of The Fertility Society of Australia.
REFERENCES
|
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media




