|
Authors: Iñaki Arroyos, María Caballero & Raquel Pillado "Up to date, clinical practice has proven that the use of in vitro culture for human embryos does not imply any major adverse consequences on the offspring" 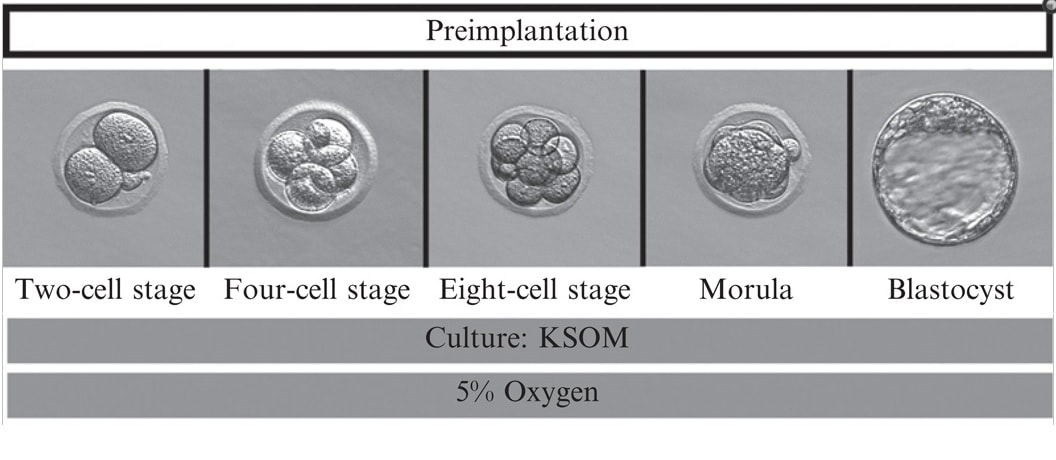 Figure 1. Mouse embryo culture from 2-cell stage to blastocyst using KSOM medium (1). Figure 1. Mouse embryo culture from 2-cell stage to blastocyst using KSOM medium (1). INTRODUCTION - Why is it important to talk about IVF culture media? Within the last 40 years, the improvement of assisted reproduction technologies (ART) has experienced rapid and great advances (2). This success is largely due to the important enhancement of the culture conditions, specially IVF media (2,3), since this is where the early embryo develops up to blastocyst stage (Figure 1) before being transferred into the womb. However, it is currently known that in spite of this huge improvement regarding culture media, there still exist certain important associated-risks. These include foetal growth restriction, premature birth, low birth weight, congenital anomalies, perinatal complications and even epigenetic alterations (2,3,4). Therefore, there is still a need to improve our current knowledge on IVF media components, concentrations and related factors, in order to dodge those potential negative effects. - How did culture media evolve through history? The development of culture media for human embryos has been possible thanks to many years of animal research (Figure 2). The first mouse embryo culture took place in plasma (5) and blood clot (6). Later on, development of synthetic media and simple culture media began, with improvements such as salt solutions supplemented with glucose, egg white and yolk (7) or Krebs-Ringer-bicarbonate (KRM) solution with glucose, bovine serum albumin (BSA) and antibiotics (8). However, embryos used at that time were collected from the Fallopian tubes at the 8-cell stage, thus being impossible to address development from the very 1-cell stage (2). Elements such as the appropriate pH level and osmotic pressure, the effect of energy substrates, BSA and amino acids in the embryo culture were widely studied during the 60’s. Glucose was replaced by lactate and pyruvate upon Brinster's demonstration that the 2-cell-stage embryo did not use glucose as energy source, and that its presence at certain concentrations hampered the 2-cell embryo development to blastocyst (10). The first success in human IVF was achieved by Edwards and Steptoe in 1969, who used a medium based on a modification of Tyrode’s solution (10). Two years later, using a more complex medium called Ham’s F-10 (made of inorganic salts, glucose, pyruvate, amino acids, vitamins and serum, among others), Steptoe and collaborators succeeded in culturing human embryos from the zygote to blastocyst (11). In 1978 and using this medium, Edwards and Steptoe got the first embryo transfer that resulted in the birth of the first IVF baby, Louise Brown (12). However, over the years, some elements from Ham's F-10 medium were shown to have adverse effects on embryo development, and so its usage was stopped (13). Years passed and different modifications took place: the removal of proteins from the media, the use of amniotic fluid and, finally, the search of the “optimal” medium. For this approach, around 1970 the development of the following media was based on the “back to nature” vision (2). New media were developed according to the composition of the reproductive (tubal and uterine) fluids (as an example, the HTF medium was made only of inorganic salts, glucose, pyruvate, lactate, human serum albumin and antibiotics). Nevertheless, glucose and phosphate were reported to be toxic to cleavage-stage embryos (as mentioned above), glutamine replaced the glucose and ethylenediaminetetraacetic acid (EDTA) was added instead of phosphate. Blastocyst formation rates were not entirely satisfactory; subsequent studies found that amino acids and vitamins improved these rates because the toxic effect of glucose was avoided (reviewed in 13). At the same time, sophisticated software was used to get a simplex optimization medium by getting the concentration of each medium component in vitro. Using mouse embryo assays, the potassium simplex optimized medium (KSMO) (14) and the KSOMAA medium (KSMO supplemented with amino acids (15)) were developed. These media are effective for the human embryo culture and, in fact, KSOMAA is widely used in human ART under the brand of Global (13). 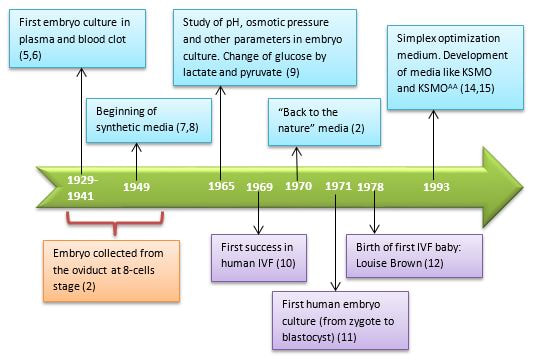 Figure 2. Overview of culture media development through history. WHICH MEDIA DO WE USE NOW? - Components Nowadays, the market of IVF culture media is widely expanded worldwide. Cook Medical, Cooper Surgical (producer of both SAGE and ORIGIO media), FertiPro, Gynotec, Genea Biomedx, InVitroCare and Kitazato are some of the companies that produce culture media for human IVF (2). Even though all commercial media present the same components (see Table 1) (2), all of them well known by every company, the concentrations and the most basic parameters for embryo culture such as the use of amino acids and glucose, temperature or gas composition, vary between brands (16). 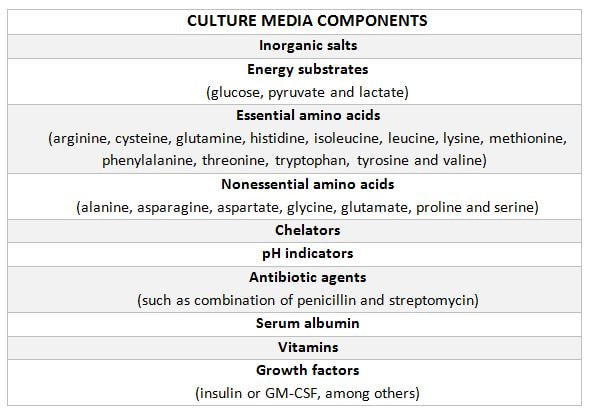 Table 1. Composition of current comercial culture media (2). - Different embryo needs (time/development stage) It is important to bear in mind the nutritional requirements of the embryo during development. Several authors have found that the required energy substrates, amino acids and proteins concentrations differed according to the embryo stage. Based on these results, it was established that between days 1 and 3 (cleavage stage), glucose should be reduced or removed if there are no amino acids or EDTA in the medium, in order to avoid the adverse effects of glucose. Lactate, pyruvate, taurine and non-essential amino acids should be included as well, but essential amino acids should be omitted at this point. A protein source such as pure (or almost pure) albumin should also be added (16). From day 3 to day 5 (blastocyst stage), the glucose concentration is increased, as well as the concentration of 20 amino acids. Pyruvate concentration in particular is lower, and protein requirements maintained (16). Change of media at day 3 is appropriate, since the embryo undergoes a series of changes at this time, such as compaction and activation of the embryonic genome (17). As a consequence, sequential media were developed. By contrast, the alternative use of single-step media is based on letting the developing embryos choose the nutrients they need. This way it is possible to minimise the stress caused by different culture environments (13,18). Both types of media are currently used by IVF clinics, even though sequential media seems to be the preferred option (18). DIFFERENCES BETWEEN CURRENT MEDIA AND TRENDS -Differences between sequential and single-step media Previously, two different kinds of media have been introduced: sequential (or two-step) and single-step (or one-step) media. In order to use these media to culture embryos from zygote to blastocyst stage, there are three different protocols available (19): [1] Sequential media protocol. Two media of different compositions are used sequentially. Medium is changed on day 3 of embryo culture. [2] Single-step protocol. Uninterrupted culture using one medium throughout the 5 days. [3] Single-step protocol with renewal. Interrupted culture using the same type of medium throughout the 5 days, including renewal on day 3. Originally, all human embryos were cultured just until day 2-3 before transfer. However, extended culture from zygote to blastocyst has attracted more attention since 1997 (18). The studies by Gardner and Lane (reviewed in 19) or Pool (16) have been the main supporters of the use of sequential media protocols. There are four main arguments in favour of the use of two-step media culture (19): [1] - The energy source required by the preimplantation embryo changes throughout development, from pyruvate and lactate to glucose, but glucose has inhibitory effects on early cleavage stages. [2] - Ethylenediaminetetraacetic acid (EDTA) helps to bypass the two-cell block. Later on, it has an inhibitory effect on blastocyst development and on the inner cell mass (ICM). [3] - Although evidence supports embryo development up to blastocyst stage with no amino acids (AA) present in the media, non-essential AAs (NEAAs), and specially glutamine (Gln) favour embryo growth during cleavage stage. Later on, during blastocyst stage, all 20 AAs stimulate the development of the inner cell mass (ICM). Furthermore, NEAAs also have an stimulation effect on the trophectoderm and the hatching process of the zona pellucida. However, an inhibitory effect on blastocyst development and viability has been observed in the presence of essential AAs (EAAs) during cleavage stage (20). [4] - The chemical breakdown of L-glutamine (Gln) in aqueous solution is the main contributor to the accumulation of ammonia during culture, which can compromise embryo development. Nonetheless, Gln is also beneficial to overcome the two-cell block. The four points mentioned above are easily addressed by using two-step media (21). However, supporters of the one-step medium protocol, in turn, argue that these problems can also be addressed through the use of one single type of medium. First, the inhibitory effects of glucose on early cleavage stages is not absolute. Furthermore, glucose is present in the natural environment of the Fallopian tubes, and there is evidence of culture media with glucose that do not inhibit embryo development (2). The reason behind this is that the inhibitory effect of glucose is determined by the interaction with other substances present in the environment. These interactions can be determined experimentally and adjustments to the concentration of medium components can be made accordingly (19). Furthermore, studies supporting the negative effects of EDTA during late developmental stages of the embryo refer to concentrations of 0.1 mmol/L; the optimal concentration needed to bypass the two-cell block is only between 0.005-0.01 mmol/L, which is insufficient to have any deleterious effect (20). Some authors like Lane and Gardner have suggested leaving out EAAs from the media during early developmental phases (19). By contrast, and after studying the AAs net depletion during human preimplantation embryo development, others such as Leese argue that the most prudent choice would be the use of a mixture with all 20 AAs. This way, the embryo would be able to choose which ones to use by itself (20). The problem of accumulated ammonium from decomposing Gln is solved by using more stable dipeptides of Gln, such as glycol-L-glutamine (GlyGln) or L-alanyl-L-glutamine (AlaGln) (19). Despite the explanation for both types of media addressing the embryo needs throughout early development, there is still a remaining question: which culture medium is more efficient? Even though in recent years multiple studies have dealt with this question, the answer is less than clear. In order to compare one-step protocols with sequential protocols, some studies compare ongoing pregnancy rates, clinical pregnancies or miscarriage rates (22). Other studies also include blastocyst formation per randomized oocyte/zygote (23), or the number and size of blastomeres in certain days, along with the final quality of the embryos (21). But the results of all of them agree in that there is no clear evidence to support either culture medium being better than the other. No significant difference was found between sequential and single medium for ongoing pregnancy rates, clinical pregnancies or miscarriage rates. Regarding embryo development, embryos cultured in one-step medium were found to present significantly more blastomeres (although unequally sized) and lower fragmentation rate in day 2 than those cultured in sequential media. However, no significant difference was observed in the percentage of good quality blastocysts between both groups (21). -Trends in the use of culture media Ever since the beginning, sequential media have been the most popular option. However, the popularity of single-step media has slowly risen within the last decade. This fact is evidenced by the increased offer of commercial media; whereas in 2008 there was only one commercial single-step medium and six sequential media (19), a few years later the number of single-step media available in the market had increased up to three times (20). Not only does the non-renewal single-step media require a reduced level of embryo handling, but it also reduces the chances of damaging or stressing the embryo. This is particularly evident in time-lapse systems, whose medium-related costs are lower. On the other hand, both the one-step medium with renewal approach and sequential protocols avoid excessive accumulation of potentially harmful waste products in the medium. Without reliable evidence supporting one type of medium over the other, it is up to the clinics to decide which one is more suitable. Therefore, further research with larger samples will be needed on comparing the efficiency of embryo culture media. CULTURE MEDIA AND PERINATAL OUTCOMES -Perinatal outcomes and assisted human reproduction Extensive animal research have provide evidence of the link between early embryo development environment and adult diseases. In the same way, in vitro culture may induce epigenetic changes in the embryo with long-term consequences, even if they are not obvious at birth or during early childhood (2). Consistently with these studies, it is known that newborns from assisted reproductive technologies (ART) often have poorer perinatal outcomes (see Table 2) compared to naturally conceived newborns. Usually, ART outcomes have been related to parental underlying medical conditions, preimplantation genetics diagnosis (if performed), cryopreservation and thawing processes, differences in hormonal treatments, laboratory conditions during embryo culture, culture media or the combination of all of them (24). 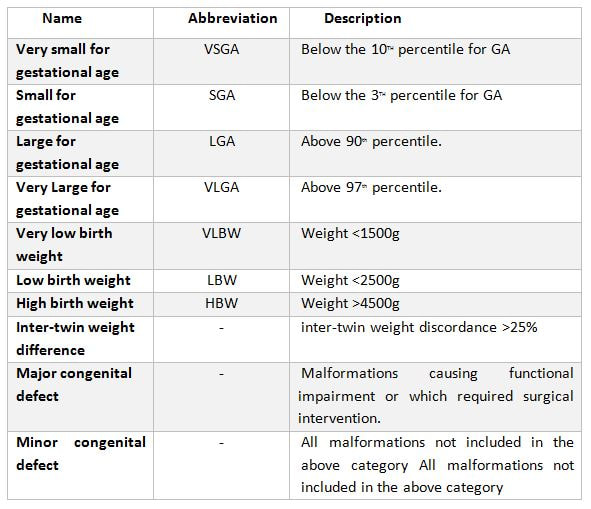 Table 2. Perinatal outcome definitions (26). Up to this day, clinical practice has proven that the use of in vitro culture for human embryos does not imply any major adverse consequences on the offspring. Nevertheless, the population born through ART is still relatively young, being Louise Brown, the first IVF child, only 49 years old. Therefore, the possible adverse repercussions on late childhood or adulthood are still a subject of study (2). -Perinatal outcomes and culture media Within last decade, multiple articles have been published that reveal actual associations between culture media and perinatal outcomes (25,26). Nevertheless, there is certain controversy when choosing between the type (single-step or sequential media) or the brand of the compared media, as well as with sample sizes. Several studies have shown no significant differences between children born after IVF and children conceived spontaneously or by intrauterine insemination (IUI) (24,25). However, there are available data that do show significant disparities in preterm birth rates between groups cultured with different media (25,26,28). Additionally, VitroLife medium seems to present a trend towards newborns large for gestational age (LGA) (25). This medium has been related to significantly higher pregnancy, clinical pregnancy and implantation rates. Simultaneously, Cook medium has been associated with lower birthweight means, higher LBW incidence and higher proportion of single embryo transfers (26). In other cases, when testing VitroLife against other media (SAGE), results indicated no significant differences for any rate; size for gestational age, LBW or even birthweight means between fresh and thawed embryos of both groups showed similar values (29). Lastly, when single-step media SSM and Global were compared, the former exhibited poorer performance than the latter, resulting in lower pregnancy and implantation rates (28). It is clear from previous studies that some culture media underperform in comparison to other media of the same type. As above-mentioned, such lower performance affects important factors like birthweight means. LGA for instance, may cause problems during labour, and LBW has been associated with higher probability of abnormal growth, neurodevelopmental problems and increased incidence of diseases such as obesity or type-2 diabetes (30). Preterm birth, in turn, may lead to the need of incubators and may cause learning disabilities or visual/hearing problems. The most likely reason behind the differences between media efficacy is the disparity in their chemical composition. For instance, whereas VitroLife uses a stable dipeptide of Gln, Cook contains L-Gln, which may be the cause for the higher percentages of LBW observed (31). APPLICATIONS AND NEW APPROACHES IN EMBRYO CULTURE - Applications Approximately, two out of three IVF cycles fail to result in pregnancy, causing significant physical, emotional and financial distress for women undergoing infertility treatments (32). In order to prevent failures, centres have chosen to perform transfer of multiple embryos, which implies a high risk of multiple pregnancy. This strategy increases the chance of developing maternal and infant morbidity; infants from multiple pregnancies are more likely to present low birthweight, and the probability of mothers suffering from miscarriage is also higher. (33). In this regard, the most common maternal complications associated with multiple pregnancies include high blood pressure, preeclampsia, increased likelihood of caesarean section, venous thromboembolism, postpartum hemorrhage and gestational diabetes (34). Considering these data, one major objective in reproductive medicine is trying to find easy, useful and clinically applicable methods to identify embryos with higher probability of implantation. Classic embryo morphology assessment is still the most established method to select embryos for transfer. However, this approach is subjected to the embryologist´s own criteria, and even though there are several scoring systems, it is not a method capable of giving reliable results (35). The recent time-lapse technology provides kinetic information of the embryos and allows the embryologist to have additional criteria for selection. But even with this improvement, more information is needed in order to find the embryo with the highest implantation potential. Nowadays, the OMICS field tries to identify biomarkers for therapeutic and diagnostic development (36). Approaches in embryo assessment based on the analysis of culture media may consider proteomics and metabolomics. Proteomics involves the study of the proteoma, including proteins secreted by embryos in the media. By studying the protein profiles, researchers are able to find altered expression of specific proteins related to important embryo events, like blastocyst development or implantation (37). The use of metabolomics, in turn, allows for the measurement of changes in the level of metabolites present in culture media that are associated with carbohydrate metabolism and amino acid turnover. The concentration of these metabolites can be used as main indicators for embryo potential. Metabolites such as glucose or lactate can effectively be associated with embryo development. Because metabolism is the final product of gene expression, the study of metabolism may be useful compared to the other OMICS, and cell function may be more accurately reflected (37). Additionally, not only metabolic flow is regulated by genetic expression, environmental stress and metabolites can be measured more precisely (39). Future directions of OMIC research should not only be focused on trying to elucidate which embryo presents the most suitable profile for implantation, but also on trying to reduce the cost of required technology and turn it affordable for daily clinical practice. - New approaches for embryo culture Culture media composition has been modified in the past recent years in order to achieve better physiological conditions. By contrast, research on embryo biophysical requirements has not developed equally fast (40). When developed in vivo, not only embryos are exposed to changing fluid chemical composition, but to mechanical stimulation (41,42,43). In addition, changes in the type of culture platform may lead to modifications in the environment immediately surrounding the embryo through regulation of chemical gradients. Thus, research on different new technologies may assist in improving embryo development (44,45). Types of culture systems
Microdrop systems have the benefit of the growth factors, with a typical drop setting of 10-50 μL. Ultramicrodrop is a variant that works with volumes of 1.5-2.0 μL, but it still presents severals typical issues related to working with small volumes (evaporation, osmolality, potential toxicity, embryo loss...) (55). A different model is conformed by the use of volumes of sub-μL (nL) of media and a vertical channel-containing polydimethylsiloxane (PDMS) culture chip (56). This allows the embryo to benefit from both reduced media volume and space. On the other hand, these same features makes the embryo difficult to recover. The Microwells system creates a microenvironment that offers a potentially increasing surface area and simultaneously a reduced space between embryos. The most popular microwell, called “well of the well” (WOW), was developed by Vajta (2000), and it has since been validated by several studies in several species (57). Initially, microwells needed to be made manually, but there currently exist available WOW systems made of polystyrene. Microchannels are related to the idea of increasing the surface area adjacent to the embryo, rather than just a single point of contact within a Petri dish. A different approach of microchannels is using glass capillary tubes filled with media (58).
Vibration systems have been developed due to the estimation that, in vivo, embryos are exposed to vibrations of 5-20 Hz in response to the ciliated epithelium of the oviduct (62). Data from different studies have shown that even short periods of gentle vibrations during oocyte maturation or early fertilization events may be of benefit to embryo quality (63,64). Finally, controlled fluid systems have been developed based on the premise that embryos, just like other cell types, can detect the shear stress (an endothelial concept) of the fluid flow (65). Excessive stress could damage the blastomeres, affect signaling pathways and cause embryo degeneration (66). CONCLUSIONS Culture media have experienced a significant development in ART, from primordial cultures like simple salt solutions supplemented with glucose and egg yolk, including changes in embryo energy sources, until more complex solutions closer to in vivo reproductive fluids. This evolution of culture media has been possible thanks to research on embryo metabolism. This allowed to find out the nutritional requirements of the embryo for every stage, thereby making it possible to develop sequential media. The development of the alternate philosophy (“let the embryo choose”) has in turn offered other advantages, such as the reduction of stress caused by different microenvironments. Both approaches have defenders and detractors, all of them with arguments to support one over the other. However, it is not yet clear which media offers the best “in vitro environment” for the embryos. Single media have gained supporters partly due to their advantages regarding reduced manipulation. This is related to time-lapse technology, even though the classical sequential media or the “renew single media” option are widely employed, too. The usage of these media is justified by the potentially deleterious effects of the accumulation of harmful waste substances. On the other hand, different ART-related procedures, including embryo culture, seem to also affect the offspring, showing poorer perinatal outcomes in comparison to naturally conceived newborns. It remains unclear whether this is derived from the culture media used or from combinations of other parameters. However, current data indicate that media do not seem to entail any major adverse consequences on the offspring. Even so, caution should be exercised. Different chemical proportions and the quality of molecular composition could explain the differences observed in some parameters like LBW between distinct commercial brands. It is important to have more information about the role of culture media in adverse perinatal outcomes. For this purpose, not just the chemical composition but also the concentrations of commercialised media should be disclosed to facilitate research on this subject. It seems obvious that modifications in the platforms used for culture media may influence embryo development. Dynamic systems are a new field of research, which may potentially help understand the physiological requirements of the embryo. However, it might result difficult to eventually improve these strategies due to extra expenses. In order to achieve so, such new devices need to count on the companies´ trust, since they are the actual responsibles for making it possible to spread technology throughout the clinical community. REFERENCES 1- Nowotschin S, Ferrer-Vaquer A, Hadjantonakis AK. Imaging Mouse Development with Confocal Time-Lapse Microscopy. Methods Ezymol. 2010;476:351-377. 2- Chronopoulou E, and Harper JC. IVF culture media: past, present and future. Hum Reprod Update. 2015;21(1):39-55. 3- Yin TL, Zhang Y, Li SJ, et al. Culture media influenced laboratory outcomes but not neonatal birth weight in assisted reproductive technology. J Huazhong Univ Sci Technolog Med Sci. 2015;35(6):932-937. 4- Sirard MA. The influence of in vitro fertilization and embryo culture on the embryo epigenetic constituents and the possible consequences in the bovine model. J Dev Orig Health Dis. 2017;8(4):411-417. 5- Lewis WH and Gregory PW. Cinematographs of living developing rabbit-eggs. Science. 1929;69:226–229. 6- Kuhl W. Untersuchungen uber die cytodynamik der furchung und fruhentwicklung des eis der weissen maus. Abb Senchenb Naturforsch Ges. 1941;456:1–17. 7- Hammond J. Recovery and culture of tubal mouse ova. Nature. 1949;163:28–28. 8- Whitten WK. Culture of tubal mouse ova. Nature. 1956;177:96–96. 9- Brinster, R.L. Studies on the development of mouse embryos in vitro. II. The effect of energy source. J. Exp. Zool. 1965;158:59–68. 10- Edwards RG, Bavister BD, Steptoe PC. Early stages of fertilization in vitro of human oocytes matured in vitro. Nature. 1969;221:632–635. 11- Steptoe PC, Edwards RG, and Purdy JM. Human blastocysts grown in culture. Nature. 1971;229:132–133. 12- Steptoe PC, and Edwards RG. Birth after the reimplantation of a human embryo. Lancet. 1978;2:366–366. 13- Yao T, Asayama Y. Human preimplantation embryo culture media: past, present, and future. J. Mamm. Ova Res. 2016;33(1):17–34. 14- Lawitts JA, and Biggers JD. Culture of preimplantation embryos. Methods Enzymol. 1993;225:153–164. 15- Ho Y, Wigglesworth K, Eppig JJ and Schultz R.M. Preimplantation development of mouse embryos in KSOM: augmentation by amino acids and analysis of gene expression. Mol. Reprod. Dev. 1995;41:232–238. 16- Pool TB. An update on embryo culture for human assisted reproductive technology: media, performance, and safety. Semin Reprod Med. 2005;23(4):309-318. 17- Quinn P. Culture systems: sequential. Methods Mol Biol. 2012;912:211-30. 18- Vajta G, Rienzi L, Cobo A, and Yovich J. Embryo culture: can we perform better than nature? Reprod Biomed Online. 2010;20(4):453-469. 19- Biggers, JD and Summers MC. Choosing a culture medium: making informed choices. Fertility and Sterility 2008, Volume 90, Issue 3, 473 – 483. 20- Gruber, I, and Matthias K, Embryo Culture Media for Human IVF: Which Possibilities Exist? Journal of the Turkish German Gynecological Association 12.2 (2011): 110–117. 21- Paternot G, Debrock S, D´Hooghe TM, and Spiessens C. Early Embryo Development in a Sequential versus Single Medium: A Randomized Study. Reproductive Biology and Endocrinology : RB&E 8 (2010): 83. 22- Dieamant, F, Petersen CG, Mauri AL, et al. Single versus Sequential Culture Medium: Which Is Better at Improving Ongoing Pregnancy Rates?, A Systematic Review and Meta-Analysis.” JBRA Assisted Reproduction 21.3, 2017: 240–246. 23- Sfontouris, IA, Martins WP, & Nastri CO, et al. Blastocyst Culture Using Single versus Sequential Media in Clinical IVF: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Journal of Assisted Reproduction and Genetics 33.10 (2016): 1261–1272. 24- Marianowski P., Dabrowski FA, Zygula A, et al. Do We Pay Enough Attention to Culture Conditions in Context of Perinatal Outcome after In Vitro Fertilization? Up-to-Date Literature Review. BioMed Research International, vol. 2016, Article ID 3285179, 6 pages, 2016. doi:10.1155/2016/3285179. 25- Dumoulin JC, Land JA, Van Montfoort AP, et al. Effect of in vitro culture of human embryos on birthweight of newborns Human Reproduction, Volume 25, Issue 3, 1 March 2010, Pages 605–612. 26- Nelissen EC, Van Montfoort AP, Coonen E, et al. Further evidence that culture media affect perinatal outcome: findings after transfer of fresh and cryopreserved embryos. Human Reproduction, Vol.27, No.7 pp. 1966–1976, 2012 doi:10.1093/humrep/des145. 27- Sandi-Monroy NL, Winkle T, Huzurudin N, et al. Does in vitro culture of human preimplantation embryos negatively affect perinatal outcomes? American Society for Reproductive Medicine Vol. 102, No. 3, Supplement, September 2014 pages 306-307. 28- Bouillon C, Léandri R, Deschet L, et al. Does Embryo Culture Medium Influence the Health and Development of Children Born after In Vitro Fertilization?. PLoS ONE 11.3 (2016): e0150857. 29- Fang Gu, Mingfen Deng, Jun Gao et al. The effects of embryo culture media on the birthweight of singletons via fresh or frozen-thawed embryo transfer: a large scale retrospective study. BMC Pregnancy and Childbirth, 2016, 16:270 DOI 10.1186/s12884-016-1077-7. 30- World Health Organization (WHO). Preterm birth. Fact sheet, November 2017 31- Zandstra H, Van Montfoort APA,and Dumoulin JCM. Does the type of culture medium used influence birthweight of children born after IVF?, Human Reproduction, Volume 30, Issue 3, 1 March 2015, Pages 530–542, https://doi.org/10.1093/humrep/deu346. 32- SART. Assisted reproductive technology success rates. National summary and fertility clinic reports. Centers for disease control, USA, 2006. 33- Lieberman B, Ali R, and Rangarajan S. Towards the elective replacement of a single embryo transfer (eSET) in the United Kingdom. Human Fertility, vol.10, no 2pp.123-127, 2007. 34- Walker MC, Murphy KE, Pan S, et al. Adverse maternal outcomes in multifetal pregnancies. American Journal of Obstetrics and Gynecology, vol.111, no.11, pp.1294-1296, 2004. 35- Rocha JC, Felipe Passalia, Felipe D. Matos, Marc P. Maserati Jr, Bruna L. Cardoso, Andrea C. Basso and Marcelo F.G. Nogueira. Methods for assessing the quality of mammalian embryos. How far we are from the gold standard?. JBRA. Assist Reprod. 2016 Jul-Sep; 20(3): 150-158. 36- Zhang-Zhi Hu, Hongzham Huang, Cathy H. Wu, Mira Jung, Anatoly Dritschilo, Anna T.Riegel and Anton Wellstein. Omics-Based on Molecular Target and Biomarker identification. Methods Mol Biol. 2011;719:547-571. 37- Domínguez F, Gadea B, Esteban FJ, Horcajadas JA, Pellicer A, Simón C. Comparative protein profile analysis of implanted vs non-implanted human blastocysts. Human Reprod. 2008; 23: 1993-2000. 38- Urbanczyk-Wochniak E, Luedemann A, Kopka J, Selbig J, Roessner- Tunali U, Willmitzer L, Fernie AR. Parallel analysis of transcript and metabolic profile. EMBO Rep 2003; 4:989-992. 39- Johnson HE, Broadhurst D, Goodacre R, Smith AR. Metabolic fingerprinting of salt tomatoes. Phytochemestry 2003; 62: 919-28. 40- Gary D. Smith, Suichi Takayama, Jason E. Swain.Rethinking in Vitro Embryo Culture: New developments in Culture Platforms and Potential to Improve Assisted Reproductive Technologies. Biology of Reproduction, Volume 86, Issue 3, 1 March 2012,62,1-10. 41- Westrom L, Mardh PA, Mecklenburg CV, Hakansson CH. Studies on ciliated epithelia of the human genital tract. II. The mucociliary wave pattern of fallopian tube epithelium. Fertil Steril 1977;28: 955 961. 42- Paltieli Y, Weichselbaum A, Hoffman N, Eibschitz I, Kam Z. Laser scattering instrument for real time in-vivo measurement of ciliary activity in human fallopian tubes. Hum Reprod 1995; 10: 1638 1641. 43- Fauci L, Dillon R. Biofluidmechanics of reproduction. Annu Rev Fluid Mech 2006;38: 371 394. 44- Swain JE, Smith GD. Advances in embryo culture platforms: novel approaches to improve preimplantation embryo development through modifications of the microenvironment. Human Reprod Update 2011; 17: 541557. 45- Smith GD, Swain JE, Bormann CL. Microfluidics for gametes, embryos, and embryonic stem cells. Semin Reprod Med 2011;29:5 14. 46- Reed ML, Woodward BJ, Swain JE. Single or group culture of mammalian embryos: the verdict of the literature. J Reprod Stem Cell Biol 2011;2:77 87. 47- Borman C, Swain J, Ni Q, Kennedy R, Smith G. Preimplantation embryo secretome identification. Fertil Steril 2006; 86:s116. 48- Katz-Jaffe MG, Schoolcraft WB, Gardner DK. Analysis of protein expression (secretome) by human and mouse preimplantation embryos. Fertil Steril 2006; 86:678 685. 49- Cortezzi SS, Garcia JS, Ferreira CR, Braga DP, Figueira RC, Jaconelli A, Jr, Souza GH, Borges E, Jr, Erbelin MN. Secretome of the preimplantation human embryo by bottom- up label- free proteomics. Anal Bioanal Chem 2011; 401:1331 1339. 50- Richter KS. The importance of growth factors for implantation embryo development and in- vitro culture. Curr Opin Obstet Gynecol 2008; 20: 292 304. 51- Lane M, Garner DK. Effect of incubation volume and embryo density on the development and viability of mouse embryos in vitro. Human Reprod 1992; 7: 558 562. 52- Donnay I, Van Langendonckt A, Auquier P, Grisart B, Vansteenbrugge A, Massip A, Dessy F. Effects of co-culture and embryo number on the in vitro development of bovine embryos. Theriogenology 1997; 47: 1549 1561. 53- Spindler RE, Crichton EG, Agca Y, Loskutoff N, Critser J, Gardner DK, Wildt DE. Improved felid embryo development by group culture is maintained with heterospecific companions. Theriogenology 2006; 66: 82 92. 54- Moessner J, Dodson WC. The quality of human embryo growth is improved when embryos are cultured in groups rather than separately. Fertil Steril 1995; 64: 1034 1035. 55- Ali J. Continuos ultra micro drop culture yields higher pregnancy and implantation rates than either large- drop culture or fresh- medium replacement. Clin Embryol (Online) 2004; 7: 1 23. 56- Melin J, Lee A, Foygel K, Leong DE, Quake SR, Yao MW. In vitro embryo culture in defined, sub-microliter volumes. Dev Dyn 2009; 238:950 955. 57- Vaita G, Peura TT, Holm P, Paldi A, Greve T, Trounson AO, Callesen H. New method for culture of zona -included or zona-free embryos: the well of the well (WOW) system. Mol Reprod Dev 2000; 55: 256 264. 58- Thouas GA, Jones GM, Trounson AO. The “GO” system - a novel method of micro culture for in vitro development of mouse zygotes to the blastocyst stage. Reproduction 2003; 126: 161 169. 59- Trimarchi JR, Liu L, Smith PJ, Keefe DL. Non invasive measurements of potassium efflux as an early indicator of cell death in mouse embryos. Biol Reprod 2000; 63:851-857. 60- Lane M, Hooper K, Gardner DK. Effect of essential amino acids on mouse embryo viability and ammonium production. J Assist Reprod Genet 2001; 18: 519 525. 61- Matsumura K, Hayashi N, Kuroda Y, Takiue C, Hirata R, Takenami M, Aoi Y, Yohshioka N, Habara T, Mukaida T, Naruse K. Improved development of mouse and human embryos using a tilting embryo culture system. Reprod Biomed Online 2010; 20: 358 364. 62- Palieli Y, Weichselbaum A, Hoffman N, Eibschitz I, Kam Z. Laser scattering instrument for real time in -vivo measurement of ciliary activity in human fallopian tubes. Hum Reprod 1995; 10: 1638 1641. 63- Mizobe Y, Yoshida M, Miyoshi K. Enhancement of cytoplasmic maturation of in vitro-matured pig oocytes by mechanical vibration.J Reprod Dev 2010; 56: 285 290. 64- Isachenko E, Maettner R, Isachenko V, Roth S, Kreienberg R, Sterzik K. Mechanical agitation during the in vitro culture of human pre-implantation embryos drastically increases the pregnancy rate. Clin Lab 2010; 56: 569 576. 65- Xie Y, Wang F, Zhong W, Puscheck E, Shen H, Rappolee DA. Shear stress induces preimplantation embryo death that is delayed by the zona pellucida and associated with stress-activated protein kinase-mediated apoptosis. Biol Reprod 2006; 75: 45 55. 66- Xie Y, Wang F, Puscheck EE, Rappolee DA. Pipetting causes shear stress and elevation of phosphorylated stress-activated protein kinase/jun kinase in preimplantation embryos. Mol Reprod Dev 2007; 74: 1287 1294.
8 Comments
Ved Prakash
12/6/2017 21:47:19
Nice and very informative article.
MARIA
12/10/2017 23:32:16
Thank you very much for your kind coment Ved!
Mavidi.simhachalam
4/25/2018 05:26:15
VeryUseful information
Raquel
4/25/2018 20:39:37
We are glad that you liked our article and thank you for your comment Madivi, we really appreciate your opinion.
prof.dr.med. g.o. ajayi
5/6/2018 17:44:11
Thanks for the well presented work.
Raquel
5/6/2018 19:01:49
Thank you for your comment Dr. Ajayi, we are delighted that you liked our article! We really appreciate you taking the time to give us your opinion.
Javier Del Rio
10/16/2018 17:02:22
Thank you for your comment. We really appreciate it! Leave a Reply. |
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media
