|
"Chromosome organization has been proposed to have potential functional implications in male fertility". Author: Lidón Carretero Vilarroig 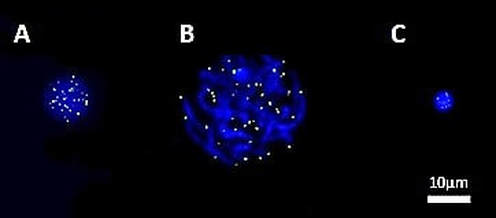 Fig. 1. FISH-PNA telomere signals (green) on different stages of sperm maturation (genetic material stained in blue): (A) spermatogonium; (B) primary spermatocyte; (C) sperm cell [in collaboration with CIMAB (Centro de Infertilidad Masculina y Análisis de Barcelona, Spain)]. WHAT ARE TELOMERES? Telomeres are nucleoprotein complexes located at both ends of linear chromosomes. In the case of vertebrates, there are hundreds of tandem repeats of the sequence TTAGGG. Together with their protein complex, telomeres play a very important role in stability and replication of chromosomes, preventing their breakage and regulating recombination between homologs (1, 2). Telomere length (TL) is determined by various loci located on several autosomes and on the X chromosome. In addition, factors such as age, gender and free radical levels have an effect on TL, as well as specifically the age of the father at the time of conception of the offspring (3). Meyerson et al estimated that the mean TL varies between 5 and 10 kb in somatic cells, and between 10 and 20 kb in germ cells, depending on the species and cell type studied (4). However, in somatic cells telomeres suffer from shortening in each round of replication; this is due to the inability of the DNA polymerase, the enzyme in charge of synthesising DNA, to elongate the 5’ end (known as “End Replication Problem”) (5). By contrast, germ cells count on an active enzyme able to solve this problem: the telomerase. Nevertheless, this enzyme is only active in specific cell lines, stem cells and immortal cancer cells. TELOMERES IN SPERM CELLS Telomeres play a main role in the organization of chromosomes inside the nucleus of sperm cells, since they take up specific non-random positions (6, 7) (Fig. 2). Although different models have been proposed to explain chromosome organization inside the sperm nucleus, it is well accepted the idea that every chromosome has a particular position, known as chromosome territory (Fig. 2) (7). Some authors have described a dynamic length during spermatogenesis: telomeres shorten during the early stages of sperm differentiation, whereas by the end of the process they elongate again, even exceeding their starting length (8). It is also known the importance of telomeres in chromosome recognition during cell division. They regulate recombination of homologous chromosomes and seem to be the first structure from the paternal nuclear genome to respond to signals after fertilization, leading to pronucleus formation (1, 2). 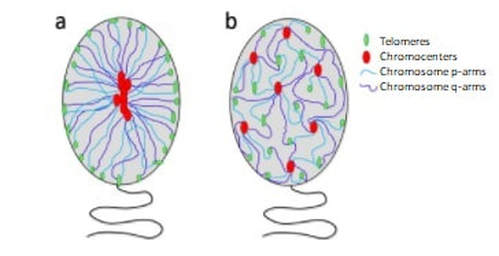 Fig. 2. Schematic representation of the proposed models of chromatin organization within sperm nuclei (7). SPERM CELL TELOMERES AND MALE INFERTILITY Telomeres have been targeted as potential candidates to explain some cases of infertility, since they are one of the epigenetic regulatory mechanisms required for proper embryogenesis (9). Due to the fact that telomeres are directly involved in chromosome recognition during cell division, modifications in the structure and length of telomeres may be related to abnormal chromosomal dotation (10). Shorter telomeres in spermatozoa may be a leading cause of aneuploidy (11), being this related to recurrent pregnancy loss. The proper organization and length of telomeres contribute to an accurate cell division, thus generating sperm cells with the right chromosome complement. Regarding their arrangement inside the nucleus, chromosome territories are thought to be organized in such a way that they allow for the actual interaction between genes from different chromosomes. Given the importance of such interactions, chromosome organization has been proposed to have potential functional implications in male fertility. Alterations in TL have been previously studied, suggesting a link between telomere shortening and errors during programmed cell death (apoptosis), sperm maturation and reduction in fertility (12, 13, 14). Also, DNA damage has been associated with disruption of the normal telomere–telomere interactions, leading to possible loss of the typical loop configuration of telomeres found in chromosomes. This would imply abnormalities in sperm chromosome deposition, activation and processing by the oocyte at the time of fertilization (15). Several studies in mice have suggested the existence of a protective mechanism that would degrade spermatocytes with reduced telomere length to prevent their maturation. Should this checkpoint fail, defective spermatocytes with shortened telomeres would be able to progress through meiosis (the typical division of gamete precursors) (16, 17), thus resulting in potentially abnormal (non-functional) sperm cells. CONCLUSIONS Telomere length could be one of the causes to explain some cases of male infertility of unknown origin. Some authors have described shortened telomeres from both sperm and ova to have additive effect on limiting the potential growth of embryos. Despite all these considerations, the actual role of telomere length in male infertility remains to be further investigated. REFERENCES 1. Ozturk, S. Telomerase Activity and Telomere Length in Male Germ Cells. Biol. Reprod. 92, 53–53 (2015). 2. Bekaert, S., Derradji, H. & Baatout, S. Telomere biology in mammalian germ cells and during development. Dev. Biol. 274, 15–30 (2004). 3. Thilagavathi, J., Venkatesh, S. & Dada, R. Telomere length in reproduction. Andrologia 45, 289–304 (2013). 4. Meyerson, M. Role of telomerase in normal and cancer cells. J Clin Oncol 18, 2626–2634 (2000). 5. Rieko Ohki, Toshiki Tsurimoto and Fuyuki Ishikawa. In Vitro Reconstitution of the End Replication Problem. Molecular and cellular biology 2001 Sept. 2001, p. 5753–5766 6. Mudrak, O., Tomilin, N. & Zalensky, A. Chromosome architecture in the decondensing human sperm nucleus. J. Cell Sci. 118, 4541–4550 (2005). 7. Ioannou, Dimitrios; Millan, Nicole M.; Jordan, Elizabeth; Tempest, Helen G. A new model of sperm nuclear architecture following assessment of the organization of centromeres and telomeres in three-dimensions. Scientific Reports. 2017/01/31/online VOL7; 41585 http://dx.doi.org/10.1038/srep41585 10.1038/srep41585 8. Tanemura, K. et al. Dynamic rearrangement of telomeres during spermatogenesis in mice. Dev. Biol. 281, 196–207 (2005). 9. Esteves SC. A clinical appraisal of the genetic basis in unexplained male infertility. Journal of Human Reproductive Sciences. 2013;6(3):176-182. doi:10.4103/0974-1208.121419. 10. Hanna, C. W. et al. Telomere length and reproductive aging. Hum. Reprod. 24, 1206–11 (2009). 11. Treff, N. R., Su, J., Taylor, D. & Scott, R. T. Telomere DNA deficiency is associated with development of human embryonic aneuploidy. PLoS Genet. 7,e1002161 (2011). 12. Fujisawa, M. et al. Telomerase activity in the testis of infertile patients with selected causes. Hum. Reprod. 13, 1476–9 (1998). 13. Cheng, E.-H. et al. Evaluation of telomere length in cumulus cells as a potential biomarker of oocyte and embryo quality. Hum. Reprod. 28, 929–36 (2013). 14. Moskovtsev, S. I., Willis, J., White, J. & Mullen, J. B. M. Disruption of telomere-telomere interactions associated with DNA damage in human spermatozoa. Syst. Biol. Reprod. Med. 56, 407–12 (2010). 15. Disruption of Telomere–Telomere Interactions Associated with DNA Damage in Human Spermatozoa. Sergey I. Moskovtsev, Jennifer Willis, John White & J. Brendan M. Mullen. Systems Biology in Reproductive Medicine Vol. 56 , Iss. 6,2010 16. Hemann MT, Rudolph KL, Strong MA, DePinho RA, Chin L, Greider CW. Telomere dysfunction triggers developmentally regulated germ cell apoptosis. Mol Biol Cell. 2001;12:2023–30. 17. Liu L, Blasco M, Trimarchi J, Keefe D. An essential role for functional telomeres in mouse germ cells during fertilization and early development. Dev Biol. 2002;249:74–84.
4 Comments
Prof premraj pushpakaran
1/7/2018 05:50:12
Prof premraj pushpakaran writes -- 2018 marks the 100th birth year of Arthur Kornberg!!!
Noemi
3/18/2018 19:30:21
Thanks for your comment, Prof premraj pushpakaran!
Noemi
3/18/2018 19:26:46
Thanks for your reply. From the team of Embryiologist Media, we are happy to read such answers so greatful. Congratulations for your baby!! Leave a Reply. |
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media
