|
Authors: Lidón Carretero Vilarroig & Belén Gómez Giménez Epigenetics in ART“Epigenetics explains why different cells perform different tasks despite their same DNA content” 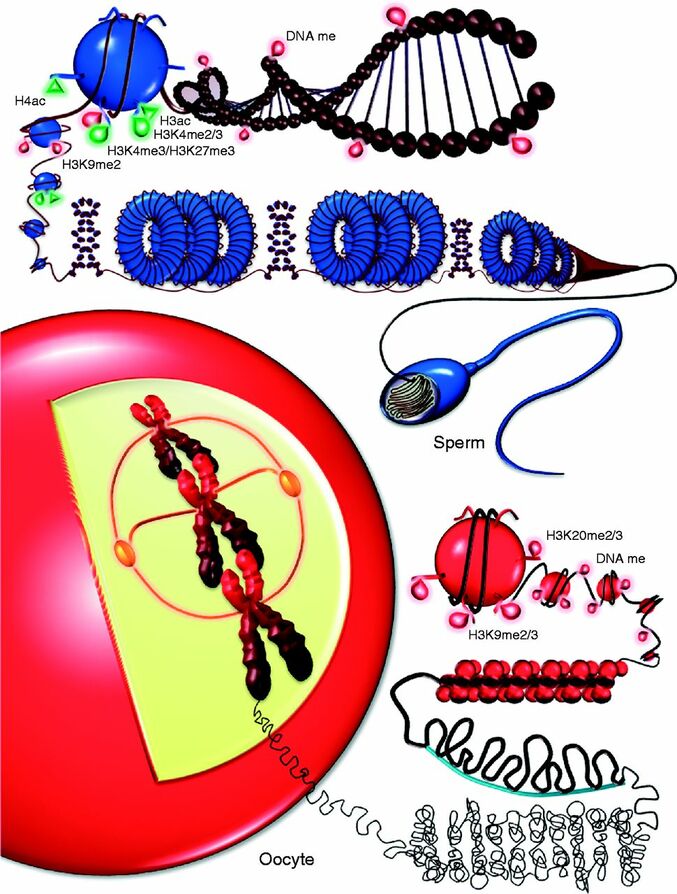 Figure 1. Epigenetic landscape in gametes (1). INTRODUCTION In vitro fertilisation (IVF) techniques have gained popularity over the years. Since the implantation of IVF techniques in 1978 more than 8 million babies have been born thanks to artificial reproductive technology (ART) worldwide (2). Nonetheless, there exists certain evidence that these techniques may induce long-term consequences on the health of the offspring (3). Consequently, exogenous hormonal stimulation, embryo culture media or embryo manipulation are currently being studied as factors affecting IVF babies. One of the mechanisms by which IVF can affect long-term health outcomes is epigenetics. This refers to modifications that affect genetic expression without altering the actual DNA sequence. The epigenome is responsible for expressing or repressing genes needed for differentiation between cellular types. Some of these epigenetic mechanisms include:
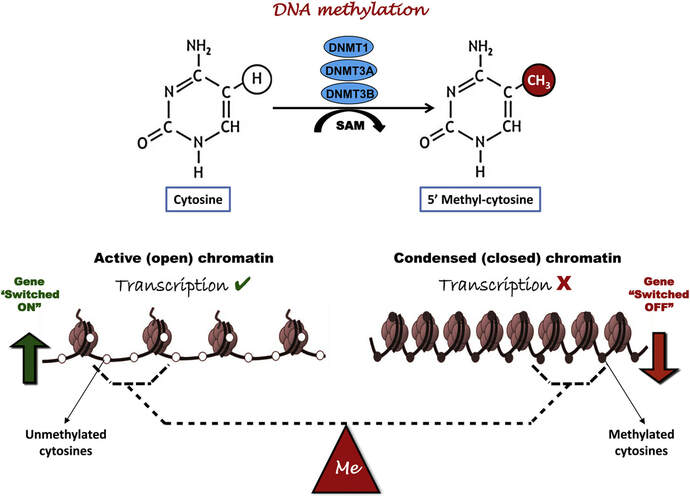 Figure 2. Methylation means the addition of a methyl group (S-adenosylmethionine (SAM)) to the C-5 position of a cytosine molecule. This transference is catalyzed by methyltransferases (DNMTs). Methylation results in chromatin condensation, thus switching off gene transcription (4). Epigenetic changes begin during gametogenesis (F1), when methylation patterns that differ between male and female gametes are established (imprinting) (Fig. 3). After fertilisation, when both gametes come together to form a zygote (F2), the embryo genome is reprogrammed, meaning every gene is de-methylated and given a new methylation pattern, with the exception of those imprinted. This is important as both gametes, oocyte and spermatozoon, are highly specialised cells with a gene expression profile suitable to their very own function. Thus, they must be reprogrammed in order to acquire pluripotency for subsequent cell division and differentiation. These changes are completed by implantation (5). Here we aim to summarize different epigenetic mechanisms in the sperm and the oocyte, as well as to review the possible effects of ART on the embryo epigenome. 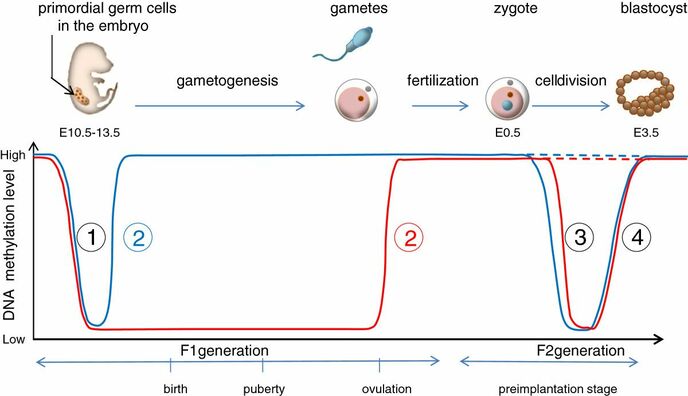 Figure 3. Reprogramming entails removal or resetting of the most previous epigenetic marks in order to allow cells to specialise and differentiate. (1) First DNA demethylation occurs in the male (blue curve) or female (red curve) primordial germ cells of F1. (2) Then, until puberty, de novo methylation takes place in the genome of the gametes. Maternal methylation marks are established at a later stage (ovulation) than paternal marks. (3) Demethylation occurs again after fertilisation in the F2 zygote. Nonetheless, paternal and maternal imprinted genes maintain their methylation pattern (dotted curves). This allows for the inheritance of parent-specific monoallelic expression in F2 somatic cells. (4) Methylation changes are completed by implantation. Modified from (5). EPIGENETICS IN THE SPERM CELL During spermiogenesis, the majority of sperm histones are substituted by protamines. Protamines are small arginine-rich proteins typical of mammals, which compact the sperm genome at a higher level than histones, thus forming a toroidal structure (Fig. 4). This structure allows for better motility of the sperm, as well as protection against exogenous DNA damage within the female tract. There are two types of protamines in humans: P1 and P2. Both work together in a 1:1 ratio in order to form the toroidal structure. Alterations of this ratio make DNA more accessible, thus increasing exposure to exogenous and endogenous sources of damage, too (6). Moreover, this imbalance is related to higher implantation and fertilisation failure rates (7). Although protamine substitution is of extreme importance for proper sperm morphology and function, about 5% of histones remain exposed to modifications. Histone-linked DNA sequences exhibit a specific pattern of modifications aimed to activate or repress marks in the promoters of genes related to imprinting and embryogenesis (8, 9). For instance, trimethylated lysine 4 on histone H3 (H3K4me3) is characteristic of developmental promoters, regions containing homeotic genes of the HOX family, some ncRNAs and paternally expressed imprinted loci (10). Alterations in some epigenetic marks such as PTMs have also been associated with fertility problems. Schon et al have recently published a study seeking alterations in histones PTMs in sperm samples (11). This study included samples from 31 men with normal or abnormal semen parameters. Their findings suggested that those with altered semen parameters displayed a decrease in histone 4 acetylation, as well as alterations in the methylation profile of H3K9 and H4K20. 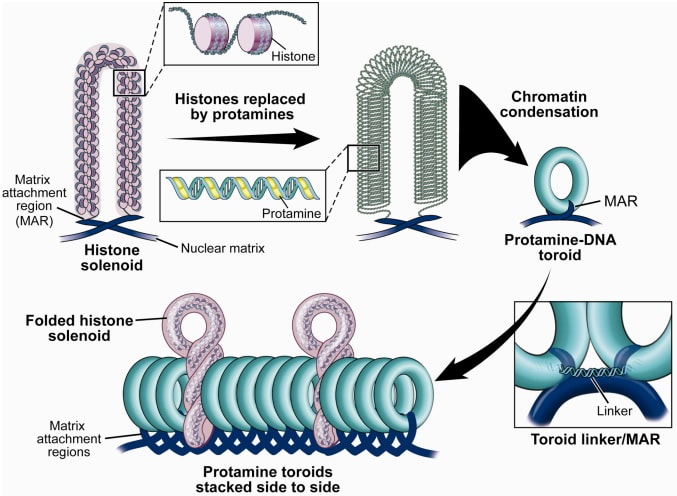 Figure 4. Sperm DNA organization. Histones are substituted by protamines P1 and P2, forming a toroidal structure. Such toroids maximize DNA compaction protecting DNA from endogenous damage. A smaller amount of DNA is kept associated with histones present in the sperm nucleus, with the remaining DNA attached to the nuclear matrix at the so-called Matrix Attachment Regions (MARs). Modified from (12). Regarding imprinting, one of the most extensively studied paternally-imprinted genes is H19. This gene is located at the long arm of chromosome 11 (11p15.5) and encodes a ncRNA involved in body weight regulation and cell multiplication (13). H19 expression is associated with another gene named Insulin Like Growth Factor 2 (IGF2). Whereas H19 acts as a tumour suppressor, IGF2 is a very important growth factor related to embryonic development, and both share common enhancers downstream of H19 (14) (Fig. 5). Demethylation of H19 activates the maternal allele, which represses IGF2; simultaneously the paternal allele is repressed because of the methylation state, thus promoting expression of IGF2 (15). Defects in H19 methylation have been detected in different cohorts of infertile men (16). Epigenetics plays an important role in fertility, but also in other phenotypic characteristics through parents to offspring. Nevertheless, the mechanisms involved in those phenotypic traits are still not well understood. PTMs are hard to study due to protamine substitution and tight packaging. For example, some authors have associated nutritional status and physical activity levels with epigenetic changes in somatic cells (17, 18), although the sperm epigenome can also become modified due to environmental factors. Recently, a link between sperm epigenome and obesity and bariatric surgery was published (19). This study showed that sperm cells from men with higher body mass index (BMI) (BMI = 31.8) have a different epigenetic profile than those with normal BMI (BMI=22.9) (19). Differences were detected in the methylation profile (methylome), as well as in the small ncRNA expression profile, whereas histone position was not altered. These results could offer an explanation as to why children from obese fathers are more likely to suffer from metabolic diseases, regardless of the mother’s weight (20, 21). 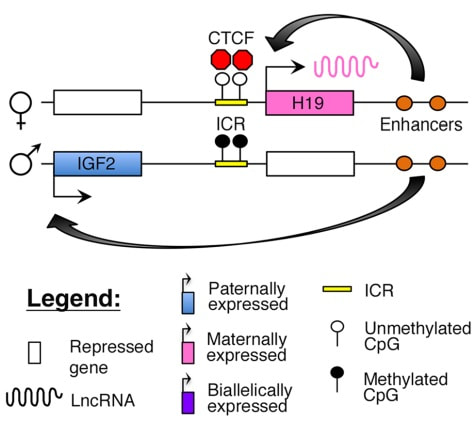 Figure 5. Proposed model of imprinting at the H19–IGF2 locus. Imprinting control region (ICR) in the H19-IGF2 locus is unmethylated in the maternal, preventing access of the IGF2 promoter, thus allowing H19 expression. On the other hand, ICR in the H19-IGF2 paternal allele is methylated, causing the repression of H19 and activation the expression of IGF2 (14). EPIGENETICS IN OOCYTES Several families of proteins contain enzymes that are responsible for maintaining the correct methylation pattern required in a particular developmental environment, such as the oocyte (22). Oocyte methylation is laid down during follicle development (1). These enzymes include, for instance, DNA methyltransferases (DNMT), demethylases (TET), histone acetyltransferases (HAT) or histone deacetylases (HDAC). As outlined above, several genes are subjected to imprinting, and their differential expression patterns depend on the epigenetic mark of each allele, which is specific for each of the parentals (23). Oocyte DNA, as opposed to sperm DNA, is compacted with histones. Research on animal models has shown that imprinted genes become methylated at different stages of folliculogenesis (24). When primordial follicles are activated, acquisition of maternal DNA methylation begins and it is completed in metaphase II (MII) oocytes (Fig. 6) (25). In mice, it is well known that maternal imprinted gene acquisition is gradual during folliculogenesis. It starts at primordial and primary follicle stages, and complete methylation occurs in the antral-ovulated follicles (24, 25). As an example, Snrpn gene is a maternally imprinted gene whose methylation begins at first stages of the follicle development. It is involved in the Prader-Willi syndrome in humans, caused if maternal methylation is not correctly established. During later stages of follicle development, PEG3 and IGFR2 genes acquire their methylation mark. Their tasks involve proliferation and intracellular trafficking. Up to early antral follicle stages, PEG1/Mest genes get their methylation pattern, playing an important role in foetal development. Finally, when follicles become antral-ovulatory follicles Impact gene becomes methylated (24, 25). This is a translational regulator that ensures constant high levels of translation upon a variety of stress conditions. Thus, similar results were observed during human oocyte growth: methylation of the SNRPN human imprinted allele starts during the germinal vesicle and metaphase I stages, and it is completed at MII stage (25). As aforementioned, the H19 gene is paternally imprinted. Its methylation is established during spermatogenesis, so the H19 allele is methylated before the maternal allele in embryos. (24). In contrast, SNRPN is methylated on both the paternal and the maternal allele but, as Lucifero et al (2004) observed, maternally imprinted alleles become methylated before the paternal ones (Fig. 6) (25). 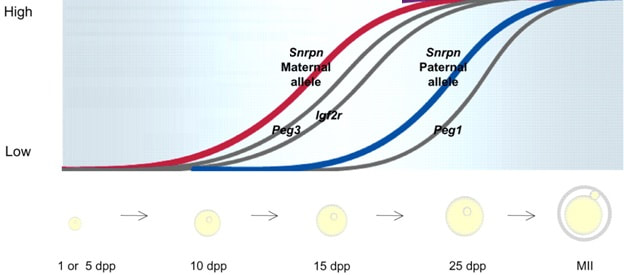 Figure 6. Schematic representation of the acquisition of the methylation profile of both paternal and maternal alleles of Peg3, Igf2r, Snrpn and Peg1 genes during oogenesis (25). EPIGENETICS IN EMBRYOS To guarantee the correct embryo genetic methylation following fertilisation, a new epigenetic reprogramming occurs again during early embryo development. Both pronuclear genomes are demethylated; by the four-cell stage the embryo genome is activated and a new increase in methylation is observed until blastocyst stage. During embryo development paternal and maternal genomes possess asymmetric epigenetic modifications (1). Uysal et al (2017) correlated demethylation and methylation patterns during oocyte maturation and early embryo development with DNMT expression (Fig. 7) (26). 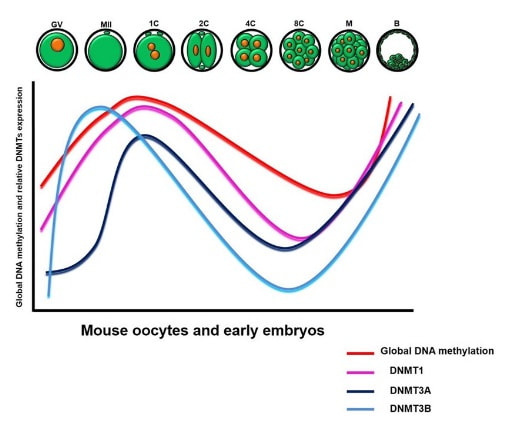 Figure 7. Global DNA methylation profile and relative expression levels of DNMT1, DNMT3A and DNMT3B proteins in the mouse oocytes and early embryos. GV: germinal vesicle oocyte, MII: metaphase II oocyte, 1C: 1-cell embryo, 2C: 2-cell embryo, 4C: 4-cell embryo, 8C: 8-cell embryo, M: morula, B: blastocyst (26). ABNORMAL EPIGENETIC CHANGES IN ART ART involves hormone stimulation, IVF or intracytoplasmic sperm injection (ICSI), embryo culture and cryopreservation among others. Even though ART is globally applied and well-established, several studies have shown association of ART with an increased incidence rate of certain imprinting disorders such as Beckwith–Wiedemann, Angelman, Prader-Willi and Russell-Silver syndromes (27, 28). ART procedures are applied during a window of important epigenetic reprogramming, gamete maturation and preimplantation embryo development (27). Furthermore, gametes and embryos are exposed to in vitro conditions, and although these aim to mimic the actual physiological conditions, it is still not clear how closely they match the in vivo environment. So, it has been suggested that these non-physiological conditions can increase the risk of developing imprinting disorders (27, 28). Several studies on animal models have suggested epigenetic changes in both gametes and embryos originated following ART. For instance, superovulation protocols have been found to result in altered levels of enzymes from the DNMT family during early embryo development in mice (26, 29). Market-Velker et al (2010) further observed demethylation of maternally imprinted genes (Snrpn, Peg3, Kcnq1ot1) and methylation of paternally imprinted genes (H19) in mouse embryos after superovulation (30). Similarly, Velker et al (2017) detected demethylation of Peg1/Mest gene during in vitro embryo culture in mice (31). Research lines focused on sheep and cattle have also observed that in vitro culture of oocytes and embryos could alter methylation and expression of imprinted genes. These observations led to the conclusion that gamete manipulation and/or ART may actually be responsible for what had been described as the large offspring syndrome (LOS), ruminant version of the Beckwith–Wiedemann syndrome (BWS) in humans, with similar phenotypical manifestations (32). As for vitrification, Ghazifard et al (2019) have recently observed increased levels of HAT and acetylation of histone 4 (acH4K12) in mouse oocytes, in comparison to fresh ones (33). These data add up to those reported by previous studies in which in vitro maturation was shown to disturb the expression of histone acetyltransferases or histone deacetylases in oocytes or embryo development (34, 35). Human studies on micromanipulation and epigenetics are hard to perform given how valuable and scarce samples are and because clinical cohort studies are extremely difficult and randomized prospective trials are impossible. Despite such obstacles, some findings have revealed an association between several imprinting disorders and ART (36). And even though studies have reported a three-fold to six-fold increase in the occurrence of BWS in association with ART when compared to the general population, abnormal DNA methylation has not been able to be consistently identified in IVF children (22). On the contrary, a recent study showed that ART manipulation after controlled ovarian stimulation does not increase the risk of abnormal expression and DNA methylation of imprinted genes (H19, SNRPN and IGF2) (37). It must be taken into consideration that some infertility problems of couples resorting to ART are related to advanced maternal age; so, it would be feasible to argue that epigenetic abnormalities may actually arise from the underlying cause of infertility rather that the treatment itself (38). Furthermore, these studies have been performed using poor-quality embryos that were unsuitable for transfer. This is why some authors have suggested that the poor quality of the samples could be the reason behind the increase of imprinting disorders and not ART itself exclusively (38). However, no clear affirmation can be stated yet regarding clear and evident causes for any of the above-summarised consequences. CONCLUSIONS The majority of the epigenetic reprogramming occurs during gametogenesis and embryo development, coinciding in time with most ART procedures. In spite of the millions of babies born through ART around the world and even though these approaches are currently considered a secure technology, several studies on both animals and humans suggest that non-physiological conditions may in fact induce aberrant epigenetic reprogramming, abnormal development and imprinting disorders. However, due to limitations in the availability of human samples, difficulty to create proper control gametes and the fact that most of these studies analyzed poor-quality embryos, a consistent relationship between an increased risk of imprinting disorders and ART exposure has not yet been properly demonstrated. Moreover, some authors suggest that a potential increase could be attributed to infertility rather than ART procedures (32, 39). Therefore, it seems reasonable to think that additional studies with non-infertile couples and good-quality embryos would be necessary to examine whether there exists an actual link between imprinting disorders and potential long-term effects of ART procedures. REFERENCES
1 Comment
|
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media
