|
Decidualization is the combination of changes on endometrium structure and hormone profiles, along with gene expression, cell differentiation and tissue modification, that allow for implantation of the embryo. Authors: Inés Abad, Roberto de la Fuente and Sara Sanz 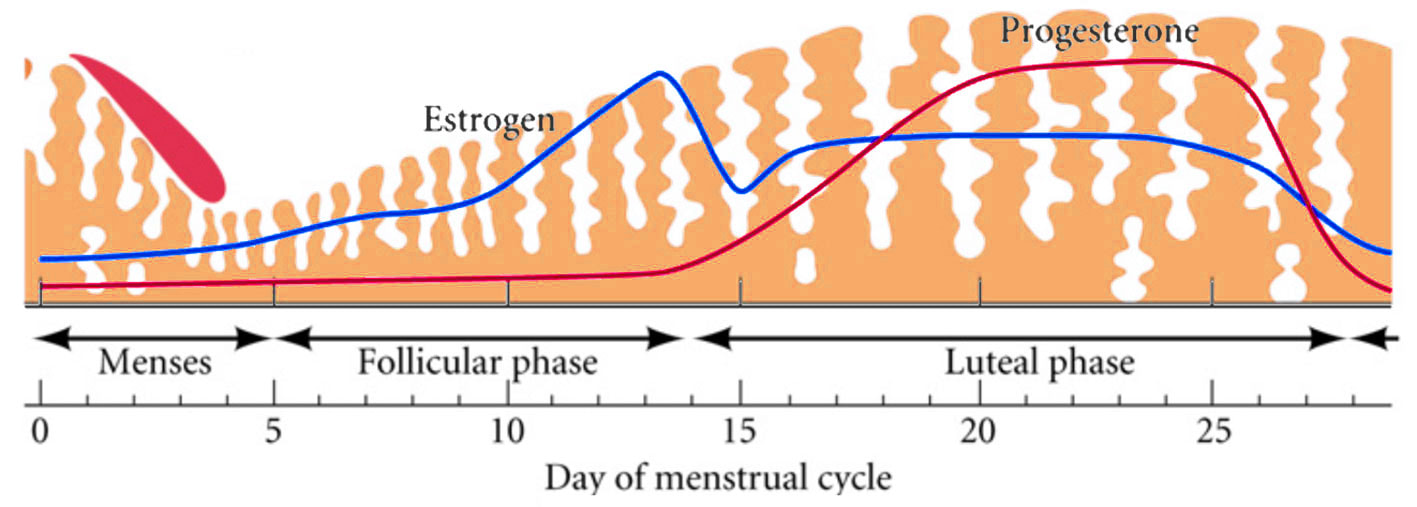 Fig. 1. Schematic overlapping view of the follicular and luteal phases of the ovary with their corresponding predominant steroid profiles and the proliferative and secretory phases of the endometrium cycle. Modified from (1). INTRODUCTION The menstrual cycle in humans lasts 28 days in regular terms, being the first day of menses considered the first day of the cycle. But in actuality, cycle refers to a series of intermingled events mainly related to three aspects: the ovary, the uterus (or specifically the endometrium) and the hormones participating in the process. As a consequence, one may refer to either the follicular or the luteal phase regarding to the ovary, the proliferative or the secretory phase of the endometrium or even to the hormone with highest blood levels. Any of them refers to a specific moment of the cycle (Fig. 2). The window of implantation (WOI) is the term used for that phase in which the uterus is ready to receive the conceptus, around day 9 after ovulation (depending on when fertilization actually occurs). In an ideal 28-day cycle, the window of implantation is considered to start at day 20 and to last for about 5 days (3). During this interval, there is a series of physiological changes in the woman (see below) aimed to facilitate the implantation of the embryo and its survival and development in the endometrial microenvironment. Because many cases of infertility have been associated with a reduced receptivity of the endometrium, a proper WOI is currently considered a fertility-determining factor (4, 5). 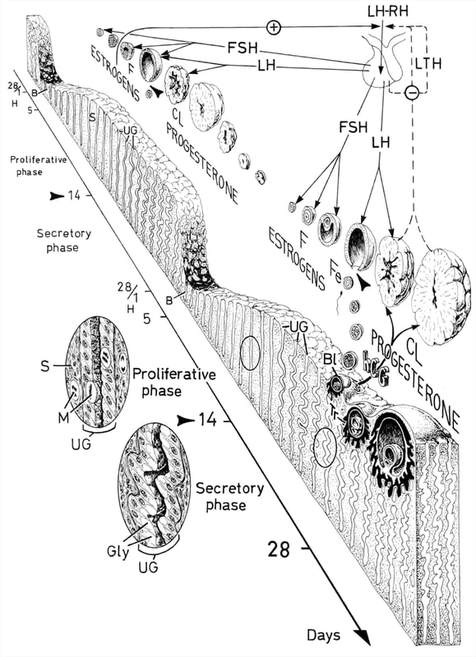 Fig 2. Illustration of the changes in the endometrium throughout the menstrual cycle in humans, along with the actions of some of the main hormones (2). Hormone fluctuation within the cycle The entire menstrual cycle is controlled by levels of different hormones, some of the most important being the gonadotropins. These hormones are glycoproteins secreted by the anterior pituitary through GnRH (gonadotropin-releasing hormone) stimulation, which is strictly controlled by the hypothalamus (reviewed in (6)). The main gonadotropins involved in regulation of the menstrual cycle are FSH (follicle-stimulating hormone) and LH (luteinizing hormone). Follicular/Proliferative phase The beginning of the cycle is regarded as the beginning of the follicular phase of the ovary and the proliferative phase of the endometrium. During this phase, cohorts of follicles at different stages progress and develop due to an increase in FSH levels. The largest follicles, called Graafian follicles (~18mm), present granulosa cells that are able to produce not only oestrogen, like early smaller follicles, but also progesterone (7). The granulosa cells are the cells that conform the follicle structure, nourishing and providing support to the oocyte. Whereas progesterone levels are relatively low and constant during the first half of the cycle, there is an increment in the blood levels of oestrogen (mainly oestradiol 17β) due to its production from the follicles. However, during the last days before ovulation, largest follicles also acquire the ability to produce progesterone, and so its levels increase significantly at that time. Oestradiol secretion creates a positive feedback that stimulates LH production; but increased levels of LH are also the result of progesterone-mediated regulation (8). Follicle growth and development means that granulosa cells not only respond to FSH-stimulation, but also to LH, as well. As a consequence, levels of oestradiol that had been increasing from ~day 5 become so high that at the preovulatory phase they trigger a rapid and highly significant release of LH from the pituitary (LH surge) (9). The LH surge causes a negative feedback on the steroidogenic pathway from the ovary, and so there is a switch from the production of oestrogen to the production of progesterone (10) (Fig. 3). Luteal/Secretory phase The LH surge coincides with ovulation (~day 14) and promotes the final stages of egg maturation to occur. At this moment, the huge amount of cells left at the follicle after ovulation form the so-called corpus luteum (CL), highly active in the production of progesterone, mainly from the granulosa cells. Oestrogen production, in turn, is back to low levels although maintained relatively constant. The rapid increase in progesterone produced by the CL has another consequence: a negative feedback that causes release of LH to cease due to inhibition of translation of the mRNA into protein, and also suppression of the growth of antral follicles due to the depression of FSH production. By the end of this phase, in the absence of conception, steroid levels are reduced at luteolysis. This results in the relaxation of the negative feedback originated by progesterone, and the restoration of positive feedback that leads to the production of oestrogens, and to FSH and LH levels reaching a plateau. As a consequence, preantral follicle development is resumed along with the next cycle (7). 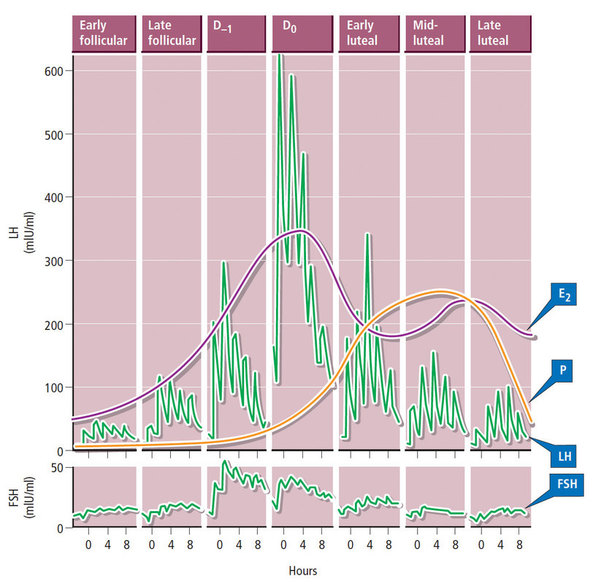 Fig 3. Profile of the main hormones involved in the menstrual cycle, with details on the different phases. Pulsatile stimulation of GnRH is seen, triggering release of LH and FSH (green profile lines). The overall levels of steroids, E2 and P, are depicted as purple and yellow lines, respectively. This image shows the clear predominance of E2 (oestrogen) in the proliferative/follicular phase vs. the main action of P (progesterone) in the secretory/luteal phase. Modified from (7). The endocrine basis of implantation It has been mentioned earlier that oestrogen (E2) is a key player during the proliferative phase of the endometrium. Besides its role in promoting stromal cell proliferation, oestrogen also induces the expression of the progesterone receptor (PR) (11), which will be of utmost importance during the secretory phase. The invasion of the endometrium by the blastocyst entails a rupture of the luminal epithelium in order to achieve implantation. For this to occur, it is necessary a series of changes in the uterine structure, as well as in the profile of several transcription factors and hormones. This way the mother adapts to the progression of the conceptus and, at the same time, controls its invasion. The development of all these changes is a process known as decidualization, which in humans starts by the mid-luteal phase, ahead of potential implantation (12). The significantly increased production of progesterone by the CL makes it the main character of the second half of the menstrual cycle. The action of progesterone has different simultaneous effects: it reduces proliferation of epithelial cells (regulated by oestrogen) while inducing their differentiation (13), and it promotes stromal cells proliferation, stimulating the glandular secretory activity. There is also a potential increase in the action of specific immune cell types (like uNKs, see below) to respond to local production of chemokines (14). Additionally, it is known that progesterone regulates the contractile activity of the junctional zone (JZ), so that it allows the conceptus to develop in a safe and controlled fashion underneath the myometrium (15). Progesterone acts partially by binding to its type A receptor (PR A) (16, 17), which in turn downregulates type B receptor (PR B), both being isoforms of the same transcriptional factor (18). The combined action of the two receptors in both stromal and glandular epithelial cells results in a modulated decidualization-like response of the endometrium in preparation for implantation. However, progesterone is not the only inducer of decidualization; in vitro progesterone only-induced differentiation of stromal cells would take up to 8-10 days under normal circumstances (19, 20). Rather, it is actually the cAMP pathway activation the main process that initiates decidualization (21, 22). In fact, many of the factors produced and secreted after ovulation like prostaglandins or gonadotropins, among others, stimulate cAMP in the stromal cells of the endometrium (22). Interestingly, cAMP levels detected from biopsies during the secretory phase are higher than during the proliferative phase (23, 24). After stimulation of cAMP, during the first stages of pregnancy, progesterone stimulates the expression of a group of interrelated genes in the endometrium, preparing the uterus for the implantation (25). Hormone profile at the beginning of gestation It has been exposed that during the luteal phase, the key acting hormone is progesterone, produced by the CL, the remnants of the Graafian follicle after ovulation. So, by the time the oocyte reaches the uterus, the endometrium is already prepared for implantation. However, if fertilization has not occurred, decidualization results useless and the physiological and endocrine profiles must be reversed. Thus, the transition from luteal to follicular phase starts around 2-3 days before the end of the menstrual cycle, with the so-called intercycle rise of FSH. This characteristic increase in FSH or FSH window is responsible for the development and eventual selection of one of the follicles in the first half of the cycle. However, after the rise of FSH, oestradiol (and inhibin B) concentration increases again, inhibiting FSH secretion, and so FSH levels are maintained (26). In humans, fertilization usually takes place in the distal third of the Fallopian tube. From there, the fertilized oocyte (now zygote) travels along the oviduct while undergoing a series of rapid divisions (cleavage), until it reaches the blastocyst stage. The blastocyst must now hatch from the hardened zona pellucida to be able to implant in the endometrium, a process that normally takes place between day 8 and 10 after ovulation (27). Not only maternal hormones regulate the process of decidualization, but the embryo itself also contributes to preparing the right microenvironment for implantation. As early as the 8-cell stage, the embryo produces hCG (human chorionic gonadotropin) (28, 29), its first active regulator of implantation. From here on, hCG secretion by the syncytiotrophoblast increases progressively after implantation, reaching detectable levels in maternal blood by day 10 of gestation (which can be normally used for pregnancy tests) and peaking around the 10th week (30). hCG has been known for decades and its current use is quite extended in reproductive medicine. In fact, it is employed to stimulate final follicle maturation before oocyte collection for IVF cycles (31), which along the external administration of progesterone for luteal phase support, helps to avoid potential problems of ovarian hyperstimulation syndrome (OHSS) derived from the use of LH. After fertilization, embryonic/placental production of hCG seems to be related with maintaining the role of the CL in secreting progesterone to sustain pregnancy (32). The need to rescue the CL from luteolysis is related to the luteoplacental shift, which occurs around the 6th week of gestation (33). This transition coincides with the decline in both progesterone and hCG serum levels detected following the 4th week and recovery thereafter (34). A crucial balance of hCG and progesterone must be reached between the CL and the syncytiotrophoblast to avoid luteal insufficiency, one of the main reasons for implantation failure during the first trimester (35). IMMUNOLOGICAL CHANGES DURING THE WOI The proliferative phase of the human endometrial cycle is characterized by the active growth of different cell types, including stromal, epithelial and vascular cells (36). On the other hand, the secretory phase supposes an increase of the glandular secretory activity and a significant increase in the number of immune cells. The major cell types in this phase are T cells, uterine NK (uNK) cells and macrophages (37). In the secretory phase, the endometrium is prepared for nidation of a conceptus, so it undergoes a series of changes during which the immune system plays an important role. Mainly, the aim of this role is to create an environment of maternal tolerance towards the conceptus, and at the same time protection against infections within the uterus (37). Immunological changes from proliferative to secretory phase T cells, essential components of the immune system and originally formed from stem cells in the bone marrow, are known to help protect the body from infections. They comprise around 45% of leukocytes in the proliferative endometrium and their number stays constant throughout the secretory phase (38). However, their relative number decreases because of a large increase in uNK cells during this phase and early pregnancy (37). During implantation, uNK cells comprise 70-80% of the total leukocyte population (39). The NK cells are a type of leukocyte or white blood cell that acts in the immune system as a first line of defense against foreign invaders. They are cytotoxic cells (40), whose function in the endometrium is to act as a source of important cytokines, including leukemia inhibitory factor (LIF), tumor necrosis factor (TNF)-α, interferon (IFN)-γ, granulocyte macrophage-colony stimulating factor (GM-CSF), and IL-10 (41). Cytokines are proteins that regulate the function of the cells that originate them on other cell types. They are primarily produced by activated lymphocytes and macrophages, and they are the agents responsible for intercellular communication, inducing the activation of specific membrane receptors, cell proliferation and differentiation, chemotaxis, growth and modulation of immunoglobulin secretion (40). Gene expression in the WOI Several genes related to the immune response are specifically regulated during the WOI. Worth to mention, among others: decay accelerating factor, indoleamine 2, 3 dioxygenase (IDO), interleukin (IL) 15, IL-15Rα, interferon regulatory factor (IRF)-1, lymphotactin, natural killer-associated transcript-2 (NKAT2) and granulysin NKG5. Some of these genes' functions are: to promote uNK cell proliferation, chemotaxis, to inhibit NK cell cytolytic activity, to inhibit cell growth and to inhibit the classical complement pathway. The functional diversity these genes represent likely serves the goal of preparing the endometrium for implantation, thereby adapting the maternal system to accommodate the immunologically ‘foreign’ fetus (42). Lobo et al. demonstrated by semi-quantitative RT-PCR (Fig. 4) and Northern analysis the upregulation of these genes in secretory versus proliferative phase (37). The mRNA of all these genes, while expressed in proliferative endometrium, demonstrated increased expression in the secretory phase. However, and taking into account the fact that uNK cells increase in number during the secretory phase, it is likely that the apparent upregulation of the above-mentioned genes is actually reflective of an increase in cell number rather than a true increase in gene expression (37). 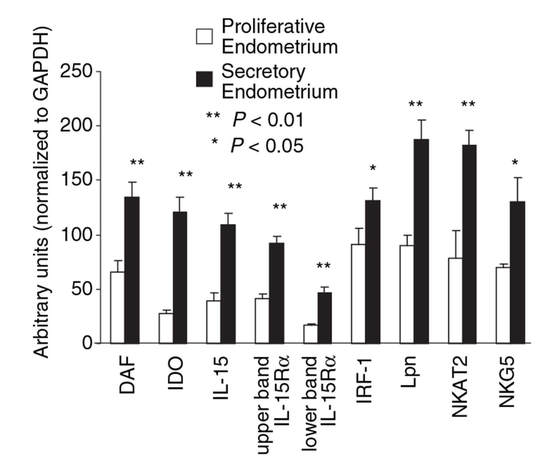 Fig.4. Graph bar showing semi-quantitative analysis of the expression of several genes, comparing proliferative (light bars) and secretory (dark bars) human endometrium. Gene expression analyzed by RT-PCR is normalized to GAPDH. There is a significant upregulation of all genes shown in the secretory phase (*P < 0.05, **P < 0.01). Modified from Lobo et al. (37). Genes that regulate an increase in uNK cell number IL-15 is a cytokine localized in the endometrium during the secretory phase of the human cycle. Its main function is to induce proliferation of uNK cells (43, 46). Some studies have demonstrated that IL-15 expression is upregulated in secretory versus proliferative phase (37, 44, 45). Additionally, Lobo et al. demonstrated the upregulation of IL-15Rα subunit from proliferative to secretory endometrium. This subunit belongs to the receptor of IL-15, and it confers high-affinity binding of cytokine to its receptor (37). IRF-1 is a transcription factor that regulates expression of IL-15 (45). IRF-1 expression has also been shown to increase during the secretory phase of the endometrium (37, 47). Genes that inhibit NK cell cytolytic activity Natural killer cells are classically considered to be cytolytic to non-self cells. Therefore, one would expect a decrease in their numbers from implantation through pregnancy to allow the fetus to be safely carried to term. Instead, there is a dramatic increase in NK cells during implantation. The answer to this paradox lies in the fact that uNK cells have effects that extend beyond their killer function, and that cytokines secreted by uNK cells are essential for implantation. In fact, NK cell-deficient mice exhibit decidual cell degeneration and endothelial cell distortion or displacement from supporting cells (48). It is believed that, at the maternal-fetal interface, human leukocyte antigen (HLA) class I molecules expressed on trophoblast cells interact with inhibitory receptors on uNK cells to protect the trophoblast from being cytolysed by uNK cells. An example of one of these inhibitory receptors is NKAT2, which shows an upregulated expression in the secretory phase (37). Genes that inhibit cell growth The uterine epithelium provides a physical barrier to infection. One mechanism for inhibiting such infections is through the secretion of endometrial peptides with bactericidal properties, which becomes especially evident at the beginning of implantation (49). NKG5 is an isoform of granulysin that exhibits antimicrobial activity against a huge spectrum of pathogens (50). The expression of this gene is especially high in NK cells surrounding the glandular epithelium in secretory endometrium (37). IDO is another example of genes that inhibit cell growth in the endometrium. It is an enzyme from the tryptophan catabolic pathway that depletes tryptophan in local tissue environments, thus suppressing proliferation of cells in the vicinity. IDO may not only inhibit T-cell proliferation, but it could also be bactericidal by this mechanism (51). An upregulation of this gene in secretory versus proliferative endometrium has been demonstrated (38). MORPHOLOGICAL CHANGES DURING IMPLANTATION WINDOW In the evaluation of women fertility, morphological studies are one of the most common procedures. These studies began with Noyes’ work, which describes the particular features of the endometrial histology, correlating it with the specific days of the menstrual cycle (52). In fact, these studies are still important and also improved and complemented thanks to the recent advances in technology. Endometrial thickness and pattern At present, there exists a data conflict between the various studies focused on endometrial thickness measurements by ultrasonography. Several papers agree in that, during an IVF cycle, the endometrium should be ≥ 7mm on the day of human chorionic gonadotropin (hCG) administration and ≥ 8mm thick on the embryo transfer day. Likewise, thickness < 6 mm has been reported to adversely affect implantation rate (53). Nevertheless, controversy arose when some studies documented no association between implantation rates and endometrial thickness. These articles claim that the important factor is the endometrial pattern, which changes throughout the entire menstrual cycle (54). Regarding endometrial pattern, the structure of a triple line is correlated with a receptive endometrium. Ultrasonography as a tool to study the endometrial pattern has a high sensitivity (79-100%), but also an elevated percentage of false positives (57-91%) (53). However, it is possible to achieve pregnancy with a “non triple-line” pattern, although at a low frequency (55). Uterine glands In the receptive endometrium, a complex secretory environment is established in order to active the blastocyst and to create both proper uterine receptivity and stromal decidualization. The structures in charge of creating this environment are the uterine glands. Once ovulation takes place, P4 levels increase and the endometrium switches into a secretory phase. The endometrium is divided into two layers, the upper one formed by loose stromal and glandular tissue in its majority, with dynamic structure and function during the cycle. On the contrary, the lower layer is structurally stable and it is formed by spiral glands and a dense stroma. During the time in which the endometrium is receptive, glycogen vesicles are disposed below the nucleus of the glandular epithelial cells, and their content is transported by microfilaments to the apical region, where glycogen is actively secreted (56). Uterine blood flow During endometrial receptivity the arterioles acquire spiral form, and there is also a similar growth of the subepithelial capillary plexus, both at basal and functional endometrium. Regarding spiral arteries, it has been reported that when glomerular capillary length reaches a certain limit, a new capillary loop is generated in order not to increase the resistance to blood flow. Additionally, during the early and mid secretory phase the vessel length, which has achieved its maximum density during the proliferative phase, experiments an increase in the number of vessel junctions (57). Recent studies have found that the existence of correct blood flow in the border area between the endometrium and myometrium plays an important role in endometrial receptivity. Uterine artery blood flow can be studied through the impedance, a term that includes the pulsatility index (PI) and the resistance index (RI) (58). Several research lines suggest that high blood flow resistance is associated with pregnant failures, whereas lower PI is a good indicator for pregnancy (Fig. 5). Applebaum et al (59). define the vascularity zones as follows:
It has been shown that pregnancy rates increase if the vessels reach the zone of the endometrium and the subendometrial halo. This fact demonstrates the relation between endometrial perfusion and endometrial growth, since the endometrium becomes thicker because of the presence of vessels. On the contrary, the absence of blood in the endometrial and subendometrial areas correlates with low probability of pregnancy, in which case uterine resistance is higher (60). Ultrasonography is also used in the study of uterine blood flow due to its non-invasiveness, real-time monitoring and repeatability. hCG administration day is normally the day for ultrasonography to be performed, because it is at this moment when highest sensitivity and specificity are achieved, although it can also be performed the day of embryo transfer (60). 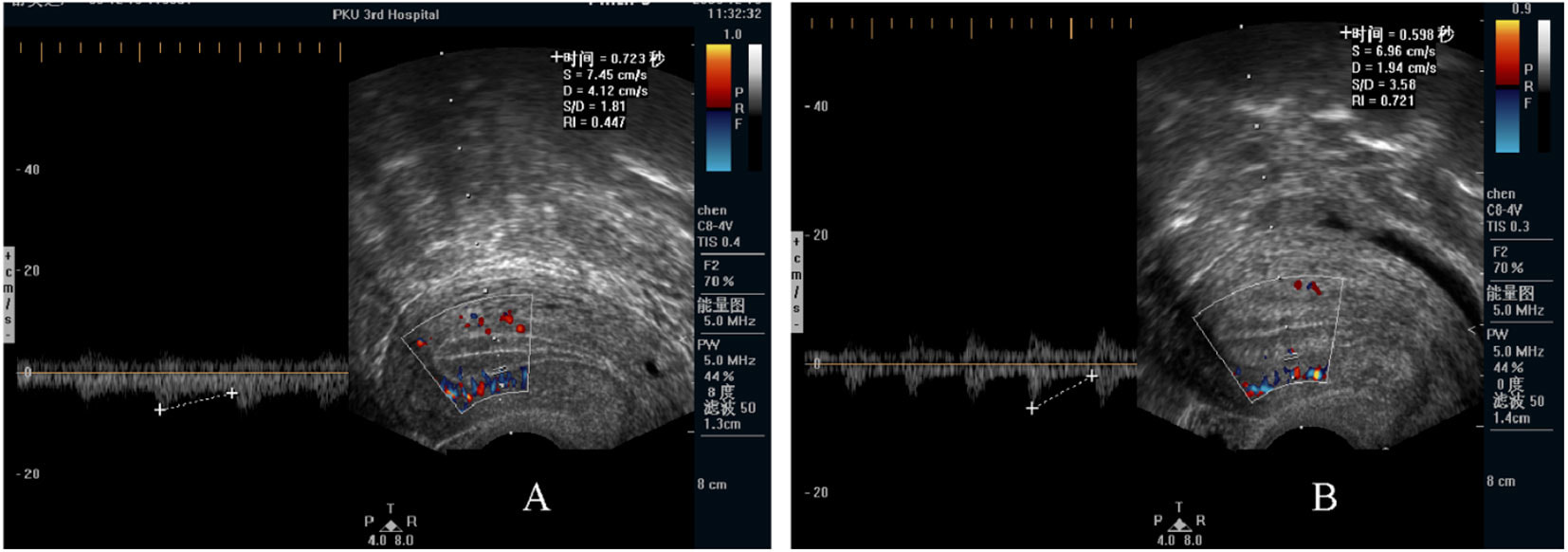 Fig 5. Endometrial blood flow. (A) Echography showing low levels of parameters such as PI, RI, S/D. (B) Endometrium of a woman with higher blood flow parameters (60). Uterine junctional zone The JZ is not only a distinct structure from the myometrium and endometrium, but it also presents functional differences (Fig. 6). There is supporting evidence for an important role of the JZ in the implantation process. On one hand, the movements of this layer allow the sperm to move rapidly to the dominant ovarian follicle. On the other hand, these contractions must decrease once fertilization has occurred. Otherwise, implantation might result impaired, reason why it is better to perform the embryo transfer during another cycle or, at least, at day 5-6 (58,61). 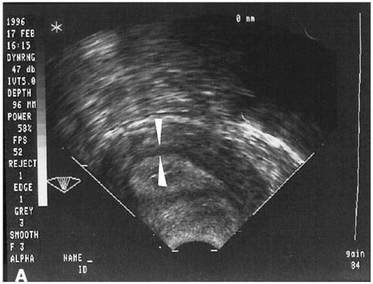 Fig 6. Echography of the uterus. Arrows show the JZ. Modified from (60). Human endometrial transcriptomics Although morphological evaluation is very useful, more advantages are needed in order to study the window of implantation. Actually, transcriptomics can be used to analyze the expression of certain genes involved in the implantation process (62). Nowadays, two existing products that can be found in the market for studying receptivity using transcriptomics are “ER Map” and “Test ERA”. These products can be used in those cases in which the uterus is apparently normal, the endometrium presents the proper thickness and yet recurrent implantation failure occurs (3 or more in case of young women and 2 or more in elderly women), especially if the embryos transferred were of good quality (63,64). CONCLUSIONS The high versatility of the endometrial structure responds to the function of housing the conceptus for implantation at the right time of the cycle. It is essential that all the endometrial features adapt to the needs of both the embryo and the mother. In order to cover its nutritional requirements and, at the same time, to regulate this extraordinary changes of the endometrium, blood supply must be increased, and so there is a significant process of angiogenesis that involves restructure of the endometrium itself. The combination of all changes needed to support early embryo implantation and further development is known as decidualization. This is triggered and controlled mainly by the action of a series of hormones, mainly progesterone from the mother and hCG from the conceptus. Production and dynamics of both hormones are in turn controlled and regulated by a network of molecular interactions, involving hormones, enzymes and other factors acting in delicate balance. This allows for adaptation to the actual and potential situations, with the goal of being always prepared during the WOI, should conception happen. The menstrual cycle exhibits extreme complexity, given the high amount of factors involved in its regulation. Even though the aspects described in the present text (hormone profiles, gene expression and physiological transformation) have been studied for decades, further research is needed to fully understand the roles of other molecular factors. Unravelling complete interaction networks of these players may uncover potential roles in implantation, thus helping find new solutions to diverse causes for infertility. REFERENCES 1. Gilbert SF, Barresi MJF. Sex Determination and Gametogenesis - The biochemistry of oocyte maturation. Developmental Biology. 11th ed: Oxford University Press; 2016 - http://11e.devbio.com/wt0610.html 2. Krstić RV. Human microscopic anatomy. An atlas for students of medicine and biology. 2nd ed. Berlin-Heidelberg: Springer-Verlag; 1994. 3. Navot D, Scott RT, Droesch K, Veeck LL, Liu HC, Rosenwaks Z. The window of embryo transfer and the efficiency of human conception in vitro. Fertil Steril. 1991;55:114-8. 4. Lessey BA, Castelbaum AJ, Sawin SW, Sun J. Integrins as markers of uterine receptivity in women with primary unexplained infertility. Fertil Steril. 1995;65:535-42. 5. Strowitzki T, Germeyer A, Popovici R, von Wolff M. The human endometrium as a fertility-determining factor. Hum Reproduction Update. 2006;12(5):617-30. 6. Knobil E, Neill JD. Knobil and Neill's Physiology of Reproduction. 4th ed: Academic Press; 2015 01 Apr 2015. 2684 p. 7. Johnson MH. Essential Reproduction: Wiley-Blackwell; 2013. 8. Dafopoulos K, Kotsovassilis CG, Milingos S, Kallitsaris A, Galazios G, Zintzaras E, et al. Changes in pituitary sensitivity to GnRH in estrogen-treated post-menopausal women: evidence that gonadotrophin surge attenuating factor plays a physiological role. Hum Reproduction. 2004;19:1985-92. 9. Kobayashi M, Nakano R, Ooshima A. Immunohistochemical localization of pituitary gonadotrophins and gonadal steroids confirms the 'two-cell, two-gonadotrophin' hypothesis of steroidogenesis in the human ovary. J Endocrinol. 1990;126(3):483-8. 10. Messinis IE. Ovarian feedback, mechanism of action and possible clinical implications. Hum Reproduction Update. 2006;12:557-71. 11. Chauchereau A, Savouret JF, Milgrom E. Control of biosynthesis and post-transcriptional modification of the progesterone receptor. Biol Reprod. 1992;46(174-177). 12. Gellersen B, Brosens IA, Brosens JJ. Decidualization of the human endometrium: mechanisms, functions, and clinical perspectives. Semin Reprod Med. 2007;25(6):445-53. 13. Weitlauf HM. Biology of implantation. In: Knobil E, Neill JD, editors. The physiology of reproduction. New York: Raven Press; 1994. p. 391-440. 14. Croy BA, van den Heuvel MJ, Borzychowski AM, Tayade C. Uterine natural killer cells: a specialized differentiation regulated by ovarian hormones. Immunol Rev. 2006;214:161-85. 15. Brosens JJ, de Souza NM, Barker FG. Uterine junctional zone: function and disease. Lancet. 1995;346:558-60. 16. Wang H, Critchley HO, Kelly RW, Shen D, Baird DT. Progesterone receptor subtype B is differentially regulated in human endometrial stroma. Mol Hum Reprod. 1998;4:407-12. 17. Bergeron C. Morphological changes and protein secretion induced by progesterone in the endometrium during the luteal phase in preparation for nidation. Hum Reprod. 2000;15((Suppl 1)):119-28. 18. Tsai MJ, O'Malley BW. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem. 1994;63:451-86. 19. Brar AK, Frank GR, Kessler CA, Cedars MI, Handwerger S. Progesterone-dependent decidualization of the human endometrium is mediated by cAMP. Endocrine. 1997;6:301-7. 20. Brosens JJ, Hayashi N, White JO. Progesterone receptor regulates decidual prolactin expression in differentiating human endometrial stromal cells. Endocrinology. 1999;140:4809-20. 21. Telgmann R, Maronde E, Taskén K, Gellersen B. Activated protein kinase A is required for differentiation-dependent transcription of the decidual prolactin gene in human endometrial stromal cells. Endocrinology. 1997;138:929-37. 22. Gellersen B, Brosens J. Cyclic AMP and progesterone receptor cross-talk in human endometrium: a decidualizing affair. J Endocrinol. 2003;178:357-72. 23. Tanaka N, Miyazaki K, Tashiro H, Mizutani H, Okamura H. Changes in adenylyl cyclase activity in human endometrium during the menstrual cycle and in human decidua during pregnancy. J Reprod Fert. 1993;97:33-9. 24. Tang B, Gurpide E. Direct effect of gonadotropins on decidualization of human endometrial stromal cells. J Steroid Biochem Mol Biol. 1993;47:115-21. 25. Bagchi IC, Cheon YP, Li Q, Bagchi MK. Progesterone receptor-regulated gene networks in implantation. Front Biosci. 2003;8:s852-s61. 26. Welt CK, Martin KA, Taylor AE, Lambert-Messerlian GM, Crowley Jr WF, Smith JA, et al. Frequency modulation of follicle-stimulating hormone (FSH) during the luteal-follicular transition: evidence for FSH control of inhibin B in normal women. J Clin Endocrinol Metab. 1997;82:2645-52. 27. Wilcox AJ, Baird DD, Weinberg CR. Time of implantation of the conceptus and loss of pregnancy. N Engl J Med. 1999;340(23):1796-9. 28. Bonduelle ML, Dodd R, Liebaers I, Van Steirtteghem A, Williamson R, Akhurst R. Chorionic gonadotrophin-β mrna, a trophoblast marker, is expressed in human 8-cell embryos derived from tripronucleate zygotes. Hum Reprod. 1988;3:909-14. 29. Lopata A, Hay DL. The potential of early human embryos to form blastocysts, hatch from their zona and secrete HCG in culture. Hum Reprod. 1989;4(8 Suppl):87-94. 30. Makrigiannakis A, Vrekoussis T, Zournakis E, Kalantaridou SN, Jeschke U. The Role of HCG in Impantation: A Mini-Review of Molecular and Clinical Evidence. Int J Mol Sci. 2017;18(6):pii: E1305. 31. Keay SD, Vatish M, Karteris E, Hillhouse EW, Randeva HS. The role of hCG in reproductive medicine. BJOG. 2004;111(11):1218-28. 32. Penzias AS. Luteal phase support. Fertil Steril. 2002;77:318-23. 33. Järvelä IY, Ruokonen A, Tekay A. Effect of rising hCG levels on the human corpus luteum during early pregnancy. Hum Reprod. 2008;23(12):2775-81. 34. Yoshimi T, Strott CA, Marshall JR, Lipsett MB. Corpus luteum function in early pregnancy. J Clin Endocrinol Metab. 1969;29:225-30. 35. Shah D, Nagarajan N. Luteal insufficiency in first trimester. Indian J Endocrinol Metab. 2013;17(1):44-9. 36. Talbi S, Hamilton AE, Vo KC, Tulac S, Overgaard MT, Dosiou C, et al. Molecular phenotyping of human endometrium distinguishes menstrual cycle phases and underlying biological processes in normo-ovulatory women. Endocrinol. 2006; 147(3): 1097-1121. 37. Lobo SC, Huang STJ, Germeyer A, Dosiou C, Vo KC, Tulac S, et al. The immune environment in human endometrium during the window of implantation. Am J Reprod Immunol. 2004; 52(4): 244-251. 38. Vassiliadou N, Bulmer JN. Quantitative analysis of T lymphocyte subsets in pregnant and nonpregnant human endometrium. Biol Reprod. 1996; 55(5): 1017-1022. 39. Pace D, Morrison L, Bulmer JN. Proliferative activity in endometrial stromal granulocytes throughout menstrual cycle and early pregnancy. Journal of clinical pathology. 1989; 42(1): 35-39. 40. Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Ann Rev Immunol. 1999; 17(1): 189-220. 41. Lash GE, Robson SC, Bulmer JN. Functional role of uterine natural killer (uNK) cells in human early pregnancy decidua. Placenta. 2010; 31: S87-S92. 42. Kao LC, Tulac S, Lobo SA, Imani B, Yang JP, Germeyer A, et al. Global gene profiling in human endometrium during the window of implantation. Endocrinol. 2002; 143(6): 2119-2138. 43. Verma S, Hiby SE, Loke YW, King A. Human decidual natural killer cells express the receptor for and respond to the cytokine interleukin 15. Biol Reprod. 2000; 62(4): 959-968. 44. Kitaya K, Yasuda J, Yagi I, Tada Y, Fushiki S, Honjo H. IL-15 expression at human endometrium and decidua. Biol Reprod. 2000; 63(3): 683-687. 45. Dunn CL, Critchley HO, Kelly RW. IL-15 regulation in human endometrial stromal cells. The Journal of Clinical Endocrinology & Metabolism. 2002; 87(4): 1898-1901. 46. Ashkar AA, Black GP, Wei Q, He H, Liang L, Head JR, et al. Assessment of requirements for IL-15 and IFN regulatory factors in uterine NK cell differentiation and function during pregnancy. The Journal of Immunology. 2003; 171(6): 2937-2944. 47. Kitaya K, Yasuda J, Fushiki S, Honjo H. Localization of interferon regulatory factor-1 in human endometrium throughout the menstrual cycle. Fertil Steril. 2001; 75(5): 992-996. 48. Greenwood JD, Minhas K, Di Santo JP, Makita M, Kiso Y, Croy BA. Ultrastructural studies of implantation sites from mice deficient in uterine natural killer cells. Placenta. 2000; 21(7): 693-702. 49. King AE, Critchley HO, Kelly RW. Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial protective role. Molecular Hum Reprod. 2000; 6(2): 191-196. 50. Ernst WA, Thoma-Uszynski S, Teitelbaum R, Ko C, Hanson DA, Clayberger C, et al. Granulysin, a T cell product, kills bacteria by altering membrane permeability. J Immunol. 2000; 165(12): 7102-7108. 51. Sedlmayr P, Blaschitz A, Wintersteiger R, Semlitsch M, Hammer A, MacKenzie CR, et al. Localization of indoleamine 2, 3-dioxygenase in human female reproductive organs and the placenta. Molec Hum Reprod. 2002; 8(4): 385-391. 52. Noyes RW, Hertig AT, Rock J. Dating the endometrial biopsy. Fertil Steril. 1950; 1:3-25. 53. Friedler S, Schenker JG, Herman A, Lewin A. The role of ultrasonography in the evaluation of endometrial receptivity following assisted reproductive treatments: a critical review. Hum Reprod Update. 1996;2(4):323–35. 54. Gingold JA, Lee JA, Rodriguez-Purata J, Whitehouse M, Sandler B, Grunfeld L, et al. Endometrial Pattern but not Endometrial Thickness Impacts Implantation Rates in Euploid Embryo Transfers. Fertil Steril. 2015;104(3):620–628.e5. 55. Racowsky C, Jackson KV, Cekleniak NA, Fox JH, Hornstein MD, Ginsburg ES. The number of eight-cell embryos is a key determinant for selecting day 3 or day 5 transfer. Fertil Steril. 2000;73(3):558–64. 56. Tapia AA. Endometrial Receptivity to Embryo Implantation: Molecular Cues from Functional Genomics. In: Violin Pereira LA, editor. Embryology - Updates and Highlights on Classic Topics: CC BY 3.0 license; 2012. 57. Gambino LS, Wreford NG, Bertram JF, Dockery P, Lederman F, Rogers PAW. Angiogenesis occurs by vessel elongation in proliferative phase human endometrium. Hum Reproduction. 2002; 17(5): 1199-206. 58. Abdallah Y, Naji O, Saso S, Pexsters A, Stalder C, Sur S, et al. Ultrasound assessment of the peri-implantation uterus: a review. Ultrasound Obstet Gynecol. 2012;39(6):612–9. 59. Sardana D, Upadhyay AJ, Deepika K, Pranesh GT, Rao KA. Correlation of subendometrial-endometrial blood flow assessment by two-dimensional power Doppler with pregnancy outcome in frozen-thawed embryo transfer cycles. J Hum Reprod Sci. 2014;7(2):130–5. 60. Wang L, Qiao J, Li R, Zhen X, Liu Z. Role of endometrial blood flow assessment with color Doppler energy in predicting pregnancy outcome of IVF-ET cycles. Reprod Biol Endocrinol RBE. 2010;8:122. 61. Lesny P, Killick SR. The junctional zone of the uterus and its contractions. BJOG Int J Obstet Gynaecol. 2004;111(11):1182–9. 62. Díaz-Gimeno P, Ruíz-Alonso M, Blesa D, Simón C. Transcriptomics of the human endometrium. Int J Dev Biol. 2014;58(2–4):127–37. 63. Endometrial Receptivity Map. iGLS: http://www.igls.net/services/service-endometrial-receptivity-map/ 64. Test de receptividad endometrial | ERA. Igenomix: http://www.igenomix.com/es/tests/test-receptividad-endometrial-era/
3 Comments
|
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media
