|
Authors: Shuyana Deba, Javier Del Río and Sara Sanz  Figure 1: MII oocytes obtained in vitro (1) Some weeks ago, Dr. Hayashi's group published a study in Nature in which they described how to generate functional mouse eggs from cultured stem cells. To better understand this experiment, it is necessary to explain the natural ovarian cycle. Although the experiment was carried out in mice, we believe these experiments could be of great importance in humans in the near future. Therefore, the following description applies to human ovarian cycle. GONADAL DEVELOPMENT IN THE EMBRYO As the yolk sac begins to incorporate into the embryo, germ cells migrate along the dorsal mesentery of the hindgut to the gonadal ridges, which they reach by the end of the fourth or early fifth week of development (2). Around week 7 of gestation, gonadal ridges undergo proliferation to form primitive sex cords, to which primordial germ cells (PGCs) are associated (Fig. 2). In the absence of both expression of the SRY gene product and male sex hormones, differentiation into female organs is determined (3,4). Whereas the origin of the follicular cells of the ovary remains unclear, likely candidates are cells from the coelomic epithelium and the mesonephros. The follicular cells eventually associate with the PGCs to form primordial ovarian follicles (3,4). ORIGIN OF GERM CELLS Human oocytes are derived from round PGCs that can be identified in the wall of the yolk sac as early as 24 days after fertilization. Once the gonad develops into an ovary, PGCs become oogonia, and mitotic divisions go on up to the seventh foetal month, and cease at some point shortly before birth (5). By the 8-9th week after fertilization, some oogonia enter into prophase I of meiosis, thus becoming primary oocytes. Meiosis starts in the deepest part of ovary (medulla) and progresses towards the cortex. Some time after it has been incorporated into a primordial follicle, the oocyte arrests after diplotene, at a particular stage called dictyate or dictyotene. This means the oocyte enters into a long quiescence period that begins before birth and ends up either resuming after the LH surge (leading to ovulation) or in follicular atresia (3). Even though the experiment by Hayashi's group has just been successfully performed in mice, the purpose of the investigation is eventually applying it to humans. 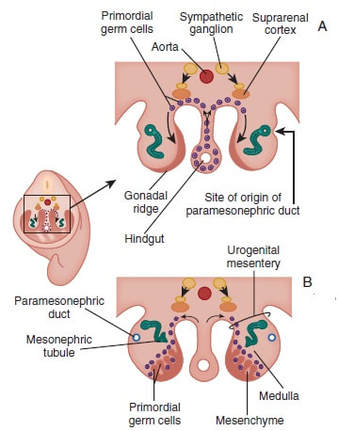 Figure 2: Gonadal development. C: Transverse sections showing the gonadal ridges and the migration of primordial germ cells into the developing gonads; D: transverse section through a 6-week-old embryo, showing the primary sex cords and the developing paramesonephric ducts (3) FOLLICULOGENESIS AFTER BIRTH Once placental oestrogen disappears, gonadotrophin levels begin to increase after birth. This increment will last 12-24 months postpartum in women. Thus, the ovaries remain inactive until the beginning of pubertal stage, moment when they will start showing response to GnRH pulsatility (6). By this time, ovaries contains germ cells at the primordial follicle phase, which are composed of small immature oocytes arrested at dictyate, and surrounded by a single layer of flat squamous granulosa cells (7). WHAT HAPPENS AFTER PUBERTY? Once sexual maturity is reached, multiple primordial follicles are activated from their quiescent state, resuming folliculogenesis by cohorts every menstrual cycle. During follicle maturation, follicles are subjected to activin, AMH, FSH, inhibin, LH, TGFβ and a multitude of other molecules that control the process. At the beginning of this process, as the oocyte resumes meiosis follicular cells proliferate and surround the oocyte, which develops into a primary follicle (7,8,9). Briefly, transition into the secondary or antral follicle stage involves the alignment of the stroma around the basal lamina and the development of an independent blood supply. Follicular cells of the underlying stroma, the granulosa cells, subsequently differentiate into an inner and an outer theca layers, the latter supporting this blood supply. The production of follicular fluid by these cells promotes the formation of the antrum in pre-antral follicles (10). Progression through antral follicle development is promoted by FSH and coincides with (7): - Completion of oocyte growth. - Acquisition of competence to complete meiosis (dependent on fertilization). - Continued granulosa cell proliferation. - Differentiation of cummulus and mural granulosa cells upon formation of the follicular antrum. Response to FSH causes growth of the follicle by proliferation of follicular cells and coalescence of the astral cavities, thus generating a follicle whose volume keeps increasing. The final stage of folliculogenesis takes place when the Graafian follicle reaches its maximum size (around 20 mm diameter). The LH surge triggers the ovulation of a metaphase II-arrested oocyte (along with the cummulus), capable of being fertilized and able to support embryonic development (7). Although there are notable differences between species, in humans, usually only one follicle is capable of completing growth and maturation up to ovulation (preovulatory), due to its more efficient response to FSH in a "competitive manner" against the rest of developing follicles. The one follicle will progress until subsequent fertilization (dominant follicle), whereas the remaining follicles in the cohort will undergo atresia (8). 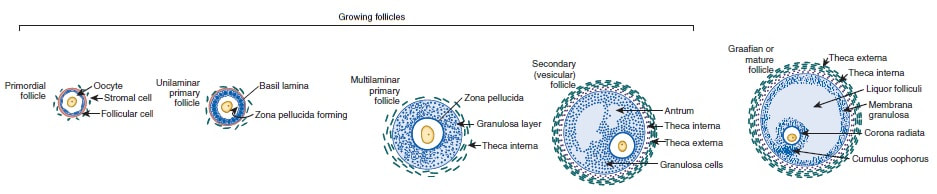 Figure 3. Shematic drawing of development of ovarian follicles (3) ABOUT PLURIPOTENT STEM CELLS For centuries, scientists have investigated how oocytes and sperm develop in mammals. Most of this work has been performed in non-human species, mainly the mouse. Human pluripotent stem cells (PSCs)-derived gamete research presents notable scientific value and potential, both for understanding basic mechanisms of gamete biology and for overcoming clinical problems (11). PSCs are currently used to answer important scientific questions such as the role of specific genes in early germ cell development, involved pathways, interaction between germ cells and supporting somatic cells, or partly or entirely in vitro PSCs-derived eggs and sperm (11). Some of the potential benefits of these new techniques could be (11): - Additional options for assisted human reproduction. - New ways to prevent and treat infertility, genetic diseases and some types of cancer. - Optimization of the number of oocytes obtained from women. "IN VITRO RECONSTITUTION OF THE ENTIRE CYCLE OF THE MOUSE FEMALE GERMLINE" (1) The authors have shown a method to reconstruct the entire process of mouse oogenesis in vitro, using embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) in order to obtain fully functional and mature oocytes. Why does this study mean so much? First of all, this is a big achievement in the field of regenerative medicine, through which it is possible to study the function of certain genes involved in oogenesis. Furthermore, it sets the ground for future similar studies and culture systems in other specie, and as such, it may result in a great advance in reproductive medicine. However, which are the limitations for this approach? The culture system requires somatic cells, which are obtained from embryos. So far, this is an ethical and legal issue in humans. A feasible solution would be to obtain the gonadal somatic cell-like cells from PSCs. The authors found that the differentially expressed genes (DEGs) were down-regulated in in vitro MII oocytes, whereas in vivo MII oocytes showed an increase in the number of transcripts. This maybe a consequence to the fact that, during in vitro growth (IVG) and in vitro maturation (IVM), oocyte development was jeopardized within a subset of (or perhaps all) the oocytes. Why? Misregulation of such genes, related to mitochondrial functions, might attenuate the potential of the in vitro MII oocytes. This problem could be solved by an analysis of the metabolic pathway, which could in turn provide information to refine both IVG and IVM. RESULTS, "PROBLEMS" AND "ACHIEVEMENTS" OF THE EXPERIMENTS (1)  As it can be seen concluded from above, it seems clear that further tests are still needed, which would help measure some aspects of the viability and functionality of this in vitro generated gametes.
REFERENCES 1. Hikabe O, Hamazaki N, Nagamatsu G, Obata Y, Hirao Y, Hamada N et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature. 2016;. 2. Baillet A, Mandon-Pepin B. Mammalian ovary differentiation–A focus on female meiosis. Molecular and Cellular Endocrinology. 2012;356(1-2):13-23. 3. Bolender DL, Kaplan S. Chapter 3 – Basic Embryology. In: Fetal and Neonatal Physiology. 2017. p. 23–39.e2. 4. Parker KL, Schimmer BP. Chapter 8 – Embryology and Genetics of the Mammalian Gonads and Ducts. In: Knobil and Neill’s Physiology of Reproduction. 2006. p. 313–36. 5. Racowsky C, Gelety TJ. Chapter 4 – The Biology of the ovary. Princ Med Biol. 1998;12:77–102. 6. David Vantman B, Margarita Vega B. Fisiología reproductiva y cambios evolutivos con la edad de la mujer. Revista Médica Clínica Las Condes. 2010;21(3):348-362. 7. Hutt K, Albertini D. An oocentric view of folliculogenesis and embryogenesis. Reproductive BioMedicine Online. 2007;14(6):758-764. 8. Green L, Shikanov A. In vitro culture methods of preantral follicles. Theriogenology. 2016;86(1):229-238. 9. Palermo R. Differential actions of FSH and LH during folliculogenesis. Reproductive BioMedicine Online. 2007;15(3):326-337. 10. Gougeon A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Annales d'Endocrinologie. 2010;71(3):132-143. 11. Mathews D, Donovan P, Harris J, Lovell-Badge R, Savulescu J, Faden R. Pluripotent Stem Cell-Derived Gametes: Truth and (Potential) Consequences. Cell Stem Cell. 2009;5(1):11-14.
1 Comment
|
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media
