|
Authors: Iris Martínez Rodero and Raquel Pillado González "Multinucleated blastomeres (MNBs) present in embryos are morphological abnormalities of unclear origin, which have been extensively correlated with chromosomal defects, lower blastocyst formation and implantation rates". 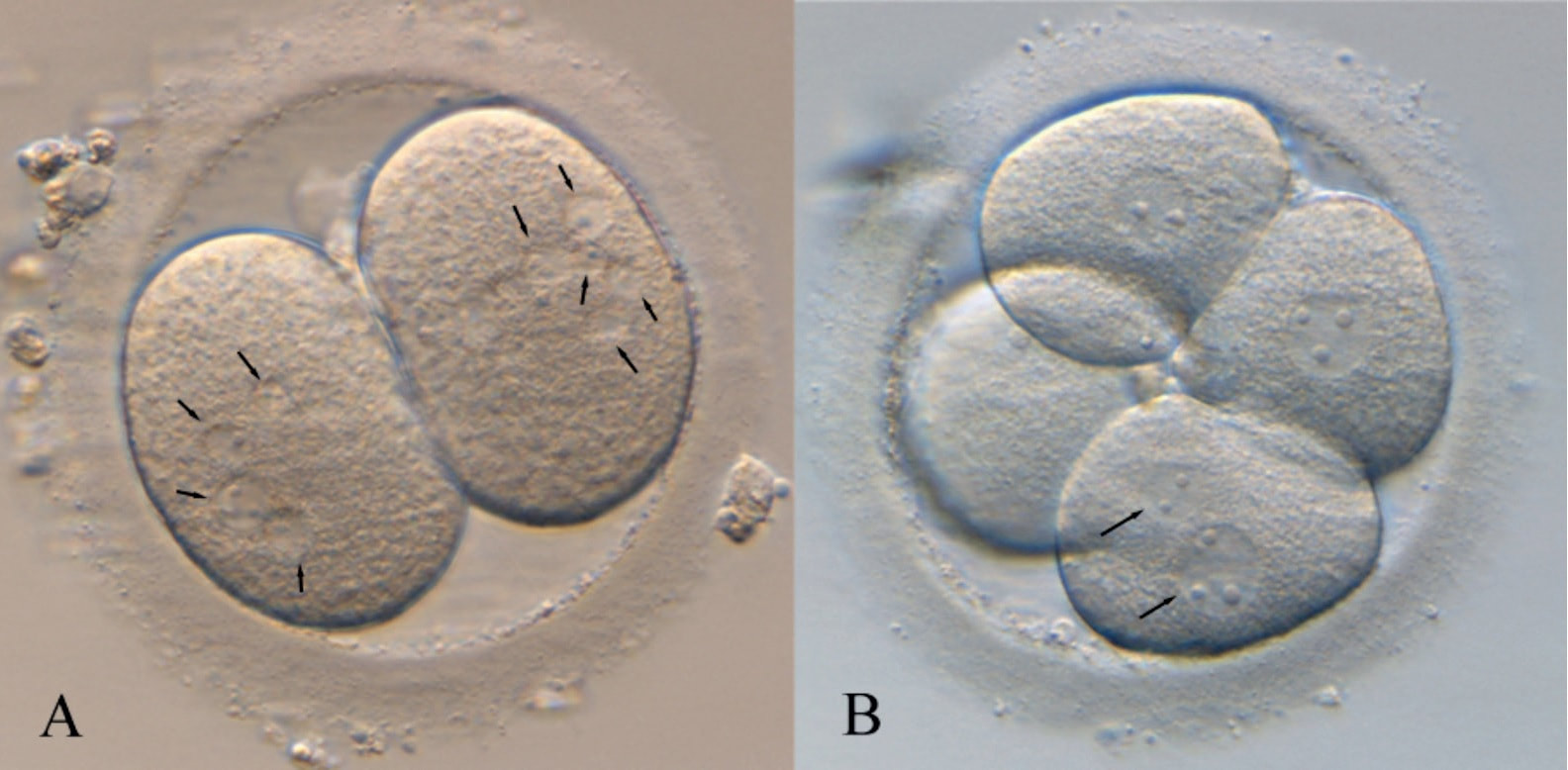 Fig 1. (A) Embryo displaying multi-/micronucleation in both blastomeres; (B) Embryo with binucleation in one of the blastomeres. Arrows point to the position of multiple nuclei (1). INTRODUCTION Selection of high-quality embryos is an important factor for the successful outcome of assisted reproduction technologies (ART). Nowadays, criteria for selection are mainly based on morphological features such as embryo fragmentation, cell number, blastomeres uniformity, etc. (2). The parameters studied so far have been demonstrated to be useful indicators of embryo quality. Their evaluation is performed through non-invasive light microscopy-based analyses, usually carried out once a day at specific time points. This approach intends to minimise the events of taking the embryos out of the incubators and exposing them to undesired harmful conditions. The presence of multinucleated blastomeres (MNBs) can be regarded as one of those indicators, and even though previous studies had already connected it to DNA abnormalities and low pregnancy rates as early as in the 90s (3,4,5) the origin of this phenomenon still remains unclear. Several possible factors seem to influence its genesis, but the specific cause for its occurrence is yet to be determined (6). Since the introduction of time-lapse imaging and monitoring technology, IVF laboratories have been able to carry out more exhaustive and continuous observations on embryo development, keeping risks at a minimum (7,8). By identifying the precise timing of specific key events of blastomere cell cycle interludes and of the embryo´s overall growth it is possible to assess its quality (7). Furthermore, time-lapse imaging and monitoring systems have facilitated the study of multinucleation (MN) in relation to its incidence in time and in the population, as well as its correlation with other morphological features and clinical variables (6,7,9). In the IVF context multinucleation is defined as the presence of two or more nuclei in one or more blastomeres. Multiples studies even differentiate between binucleated and multi-/micro-nucleated (three or more nuclei) blastomeres (Fig 1) (1,4,7). Reports on this phenomenon range from 17% to 69% of the total of cultured embryos, depending on the groups used within assays and authors (10). Factors that may influence MNB appearance are numerous and have been repeatedly studied (6,7,9,11). To date, several explanations have been proposed for MNBs: dysfunction of the mitotic spindle or the occurrence of karyokinesis without cytokinesis (12); DNA breaks or imperfect mitosis (13); nuclear membrane alterations (14) and even other factors not directly responsible for MNBs and yet linked to its presence (1). FACTORS THAT MAY RELATE TO THE APPEARANCE OF MULTINUCLEATION
Results from various studies have shown a difference in the percentage of multinucleated embryos between groups that had been fertilised by traditional IVF vs ICSI. Van Royen and colleagues showed 32.7% of MN embryo in the IVF group, compared to 34.5% in the ICSI group (6). Accordingly, Walmsley et al. 2003 reported 17,2% vs 18,3% of MN in embryos derived from IVF and ICSI, respectively (11).
Apparently, the type of infertility factor seems not to affect MN rates. Some studies have reported no significant differences in the percentages of MN embryos between cases of female factor-only infertility and male factor-only infertility (32.7% and 34.7%, respectively). In addition, differences were not found between these cases and those with both partners affected by some sort of infertility, either (6). However, different studies support the relevance of oocyte culture, specially regarding certain processes that occur naturally in in vivo conditions and that are essential for the proper embryo development. Data show that both oocytes subjected to negative conditions when cultured in vivo (like stern hypoxia, for instance) and oocytes cultured in vitro derive in a higher percentage of MN embryos (15,16). Regarding male infertility, results are more controversial; whereas some studies reflect a higher MN rate for cases in which male factor is especially severe (normally derived for ICSI rather than IVF) (11), other authors show no significant differences (6).
Records exhibit that cycles with an accelerated ovulation induction response present increased MN rates (6,17). Furthermore, several studies have reported that embryos derived from patients from whom ten or more oocytes had been collected presented a significantly higher MN rate than embryos from groups of nine or fewer oocytes (6,8,18). This is in accordance with the fact that patients who need high doses of FSH present higher MN rates (6). Very short cycles and cases where high doses of GnRH are needed trigger the development of high numbers of premature follicles that produce oocytes, which despite being able to reach metaphase II and become fertilized fail to go through proper nuclear cleavage (6,18). Lastly, even though differences in GnRH doses have been associated to significant differences in the incidence of MN embryos, similar results have not been observed when using different hormones (like rFSH, r-hFSH, purified urinary FSH or urinary gonadotropin, for instance) (6).
Multiple studies have been conducted on patients ranging from 25 to 45 years old. Several authors divided data in 5-year interval groups in order to verify whether patient age correlates with MN. However, the only significant difference was found when comparing women of +40 with younger ones of -35, presenting higher degree of MN in the first group (6,9).
Chromosome polymorphisms consist in heterochromatin variability. These are usually located in the long arms of chromosomes 1, 9 and 16, and the short arms of chromosomes from the groups D and G (13, 15 and 21, 22 and Y) (19). Even though such polymorphisms are generally regarded as normal within karyotypes (20), studies indicate that some of them might be associated with certain clinical problems such as abnormal spermatogenesis (21), infertility (22,23), recurrent miscarriages (24,25) and higher rate of chromosome abnormalities among blastomeres at the cleavage stage (26,27). Sun and collaborators hypothesized that couples with chromosome polymorphisms might experience a higher rate of embryo multinucleation (19). Nevertheless, the authors found no association between chromosome polymorphisms and MN embryo formation in couples undergoing IVF (19).
- Cellular fragmentation Although MN may appear regardless of the cellular fragmentation levels, several papers support the correlation between these two features (6,28,29). In particular, Van Royen et al. divided the level of fragmentation in three categories: F1 (≤10%), F2 (10-20%) and F3 (20-30%); this study presented evidence for higher MN in F2 and F3 when compared to F1, but similar to each other (6). - Cleavage rate When 3-cell and 5‐cell day-2 embryos were observed under the microscope, both types exhibited significantly higher multinucleation (28.2% -50%) than regular ones with the ideal 4-cell cleavage pattern (with only 16.8% MN). Similarly, day-3 embryos with the typical 8‐cell stage showed significantly lower multinucleation (15.5%) than 7‐cell and 9‐cell embryos (6). As it has been mentioned, ideal 4-cell and 8-cell stages show similar MN percentages. However, application of time-lapse imaging has revealed a significant decrease in MN from the 2-cell to the 4-cell stage (from 43.2% to 15.0%). The analysis of MN in 2-cell embryos indicated that, after cleavage, the majority (52%) of 2-cell MN embryos became mononucleated, whereas only a lower percentage (34%) showed MNBs, and about 14% were of poor quality (with only one or no visible nucleus at all) (9). This decrease in the MN rate suggests that 2-cell MN embryos are able to self-correct their nuclear abnormalities. But this repair mechanism has been observed in both euploid and aneuploid embryos, therefore it cannot be used as an indicator of chromosomal normality during embryo selection (8,9,30). An extended duration of both 2-cell and 4-cell stages has been proposed as a possible indicator of the occurrence of nuclear self-correction (9). INCIDENCE OF MULTINUCLEATION IN CLINICAL IVF As previously exposed, it is through time-lapse imaging that a far higher percentage of multinucleation (25%) has been detected compared to static observations on day 2 at 42 hours post-insemination (hpi) (<5%) (7). These observations have demonstrated that multinucleation is a frequent event that, according to Yilmaz et al., is present in at least one embryo in 41.3% of IVF cycles (31). Data provided by Desai and colleagues reported that approximately 56% of binucleated embryos and 48% of those with three or more nuclei went on to form blastocysts that met the appropriate criteria for vitrification (7). In addition, data from different studies point to binucleation being more frequent than blastomeres with 3 or more nuclei (7,31,32). At the same time, multinucleation has provided an additional criterion for embryo selection, since it is mainly observed in those of poor quality and is associated with direct and/or reverse cleavage (7). It has been observed that, out of all embryos found showing direct and/or reverse cleavage, at least one fourth were also multinucleated (7). By using time-lapse, multinucleation has been repeatedly observed to be a reversible event in a high proportion of embryos (7,32). Multinucleation reversibility has been reported to be as high as 73.4% (32); this has been calculated as the proportion of embryos in which multinucleation was detected at 2-cell stage, but not visible at 4-cell stage (likely due to self-correction mechanisms, as above-mentioned). In fact, Aguilar and collaborators reported 127 multinucleated embryos at 4-cell stage out of the 479 ones initially observed to present this feature at 2-cell stage. De novo multinucleation at the 4-cell stage in turn was observed in 36 embryos (32). IMPACT OF MULTINUCLEATION ON IVF OUTCOMES Multinucleation has traditionally been related to both low blastocyst formation (33) and implantation rates (5,6,17,28,34), and linked to the likely presence of chromosome abnormalities, which consequently results in embryo arrest (35). Nevertheless and despite all the existing evidences, there is still much controversy regarding multinucleation; reports have been published revealing cases in which fully binucleated 4-cell stage embryos had eventually developed into euploid blastocysts and genetically normal children (31,34).
Although some preimplantation genetic testing (PGT) studies have shown that not all multinucleated embryos are chromosomally abnormal (31,32,36) multinucleation is predominantly associated to chromosomal defects and poor implantation prognosis (3,31,37). Kligman and colleagues published that 74.5% of multinucleated embryos were chromosomally abnormal, compared to 32.3% of non-multinucleated embryos (3). Years later, Ambroggio et al. revealed an increased incidence of aneuploidy of MN 4-cell stage embryos when compared to single-nucleated embryos (85% vs 78%), suggesting that multinucleated embryos should not be recommended for transfer in IVF cycles (37). These results were confirmed when, from 395 MN embryos tested for PGT, Yilmaz et al. reported that 82.5% of MN blastomeres exhibited two nuclei, whereas the remaining blastomeres presented a single or three or more nuclei (31). Noteworthy, binucleated patterns of multinucleation may be less detrimental, since a high percentage of embryos with such feature are euploid, compared to embryos exhibiting three or more nuclei in a single blastomere (38).
Embryo morphokinetics was studied and related to the multinucleation status in a study conducted by Meseguer’s team (32). In the study, 53.4% of a total 1676 embryos included were MN. Based upon the reported data, differences in morphokinetics between multinucleated and non-multinucleated embryos at both 2-cell and 4-cell stages comprise cleavage events involving the completion of the first mitosis and the length of the S-phase. These differences affected the following parameters: t2, t3, second cell cycle (cc2=t3-t2), t4, t6, t7 and t8. These results allowed to conclude that, if multinucleation remains at 4-cell stage, it takes longer for the embryos to complete the next cell cycle (cc3=t5-t3). Should this be true, the restoration system would not be efficient if MNBs were still observed after the 2-cell stage (32). The origin of the multinucleation phenotype has been suggested to be multiple: disruption of intracellular restructuring, remodelling or imprinting in the developing oocyte, or even alterations in DNA replication, cytokinesis or compaction during the first cell cycle (16). If multinucleation appears as a result of defects in cell function, differences in morphokinetics between MN and non-MN embryos during these early stages may be expected (32).
Opinions on the impact of the multinucleation phenotype on implantation rates diverge from each other: On one hand, cell stage for MN appearance has been proposed to exert the highest effect on the implantation rate. Authors supporting this claim are divided into two positions: those who affirm that the presence of MN at the 2-cell stage is actually insignificant in terms of differences on implantation rates, but it is at 4-cell stage when it does have a measurable negative effect (32); and the authors who argue implantation rates to be significantly reduced when MN is already observed at the 2-cell stage (8). On the other hand, the school of Meriano and coauthors affirm that binucleation is less harmful than any other type of multinucleation (16). However, Aguilar and colleagues explained that their differences with Meriano were found on the frequency of image acquisition and the systems used to measure multinucleation (32); whereas the former acquired one picture in seven different focal planes every 20 minutes, the latter recorded images every 2.5 minutes (32). In any case, it has been demonstrated that patterns of multinucleation at 4-cell stage are correlated with low implantation rates, while any of the other cases has been reported to decrease the chances to achieve pregnancy (16,32). CONCLUSIONS Multinucleation is a common and reversible event observed in human IVF embryos, and it is specially frequent as binucleation at two-cell stage. It is associated with chromosomal defects and altered morphokinetic parameters, eventhough binucleation patterns seem to be the less severe. Regarding multinucleation impact on implantation rate, results are controverted. It seems that implantation rates are not affected when multinucleation appears as two nuclei in two-cell stage. Although the presence of multinucleated blastomeres in human embryos has been associated with the above-mentioned undesired characteristics in IVF embryos, the reasons explaining its appearance and occurrence in time and its relationship with patient specifications have not been deeply studied until time-lapse systems became available. Even though different causes have been suspected to lie behind MNB development, none of them have been proved actually represent the main responsible. Nevertheless, a growing number of studies provide data untangling the relationship between MN and assisted reproduction fertilization methods (IVF and ICSI), stimulation cycles, infertility factors, culture conditions and other embryonic morphological characteristics. Even though sometimes results from different studies may seem contradictory, this might be accredited to the differences in sample sizes. All the above said, it seems reasonable to highlight the need for further research on this issue. It would be highly helpful to unveil the actual triggers of multinucleation, to develop optimal ART practices that avoid increasing MN incidence, and to unravel any other correlation with adverse embryo features during development. Deeper knowledge would help improving embryo assessment methods and, consequently, increase the rates of successful ART outcomes. REFERENCES
0 Comments
Authors: Javier del Río, Belén Gómez-Giménez & Iris Martínez "The relationship between blastocyst morphology and implantation has been investigated mainly according to expansion, trophectoderm and inner cell mass" 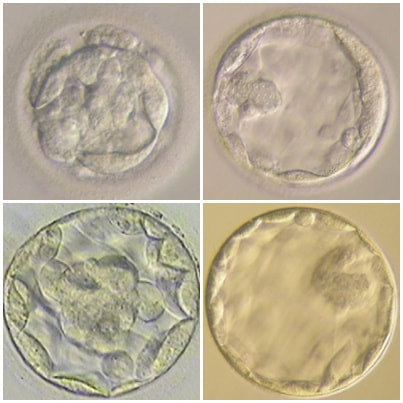 Figure 1. Human blastocysts (10). The main goal of in vitro fertilization (IVF) is the birth of a single healthy child. However, the consequences and the effects of assisted reproductive techniques on children’s short- and long-term health have always been a source of discussion. Although IVF techniques and protocols have dramatically improved, the overall success rates are still relatively low, and assisted reproduction units still face the challenge of improving pregnancy rates (1). For this purpose, transfer of a single human embryo at blastocyst stage is becoming more common in the practice of assisted reproduction (2). It allows a better synchronization between the endometrium and the embryo and the possible selection of embryos with a higher implantation potential (3). Several morphology- or kinetics-based approaches have been described to select the best blastocyst in order to increase pregnancy rates. However, the yielded results are conflicting and the outcome is a matter of never-ending and controversial debates, specially regarding blastocyst stage (4, 5). BLASTOCYST ASSESSMENT SYSTEMS The relationship between blastocyst morphology and subsequent blastocyst implantation has been investigated according to various criteria. Traditionally, morphology has been evaluated after embryo compaction (6). The significance of examining the embryo after compaction is the ability to examine it after embryonic genome activation. Furthermore, the obvious benefit of looking at the blastocyst is the possibility to examine both cell types. The extent to which the trophectoderm (TE) develops will reflect the embryo’s ability to attach and implant in the endometrium, whereas development of the inner-cell mass (ICM) is obviously crucial for the progress of the foetus (7). There have been described several assessment systems to predict the success of blastocyst implantation. However, Gardner's grading system seems to be a better predictor of pregnancy rates (8, 4, 7). Following this method, blastocysts are initially scored from 1 to 6 based on their degree of expansion and hatching status, and ICM and TE grading is then assessed from A to D depending on their morphology (9). It was felt that expansion was important for cavity formation. This process requires both extensive energy utilization through the Na+/K+ ATPases on the basolateral membrane of the TE and formation of effective tight junctions between TE cells to form a barrier. Therefore, expansion seems to be a reflection of embryo competence (7). Recently, Richardson et al. proposed a simplified blastocyst grading system. These authors demonstrated both its prognostic potential and the inter- and intra-observer variability. This grading scheme was able to effectively predict clinical outcomes in terms of implantation, clinical pregnancy and live birth. Slight variation existed both between and within embryologists grading the embryos but, overall, consistency in their analyses was similar to, if not better than, those associated with more complex grading systems (10). However, most of the grading systems that are currently used for assessing viability of IVF embryos are subjective, relying on visual inspection of morphological characteristics of the embryos that are qualitatively evaluated. Grading based on qualitative criteria is imprecise, and it inevitably results in inter-observer variability and in intra-observer to some extent, as well (10). MORPHOLOGICAL ASSESSMENT AND OUTCOME RESULTS As it has been exposed, there is a need for increased knowledge about the relative impact of each morphology parameter at the blastocyst stage (and their potential correlation) on predicting the probability of successful implantation and pregnancy (1, 2, 11). Shapiro et al. compared up to 25 parameters in order to develop predictive models of clinical pregnancy within a set of blastocyst transfer cycles (12). Among these variables, blastocyst diameter seemed to be the most significant predictor of clinical pregnancy in the multivariate models. The authors concluded that embryos developing into expanded blastocyst stage on day 5 were approximately twice as likely to implant, compared to those for which expansion was delayed until day 6 (13, 3). This is supported by Van den Abbeel and coauthors, who found that high scores of blastocyst expansion and hatching stage, ICM and TE grade were all significantly associated with increased pregnancy and live birth rates after fresh transfers (11). The finding that the expansion and hatching stage is the most important parameter when selecting a blastocyst for transfer (11) is in contrast with some retrospective cohort studies that suggest TE grading to have the strongest predictive power for treatment outcome in fresh transfers (14, 15). On the contrary, Basak Balaban et al. exposed that quantitative measurement of blastocysts and ICM is not a practical way to assess blastocyst quality, arguing that two-dimensional measurements of three-dimensional global structures can be misleading. The reasoning is that the size of a blastocyst may vary depending on the time the blastocyst is assessed under the microscope, and this may easily confuse grading (9). For this purpose, Almagor et al. tried to provide an easily measurable assessment of the ICM and evaluate its correlation with pregnancy rates in a series of single blastocyst transfers. They found a high ICM/blastocyst ratio associated with significantly increased pregnancy rates. Thus, they proposed this measure to be used as an additional strongly predictive parameter of successful implantation (16). Recently, Bouillon et al. have confirmed that clinical pregnancy and live birth rates were significantly higher for blastocysts with good TE and ICM quality, and so it was concluded that both rates decreased with morphology (4). Even though some blastocysts with non-optimal morphology are able to implant, it has been suggested that when selection is made among suboptimal blastocysts, preference should be given to those with a normal ICM (6). However, the current goal for researchers is to establish the optimal perinatal outcome of singletons according to blastocyst morphology. This has been recently analyzed by Bouillon et al., who found no increased rates of adverse obstetric and perinatal outcomes after transfer of blastocysts with poor morphological features (4). 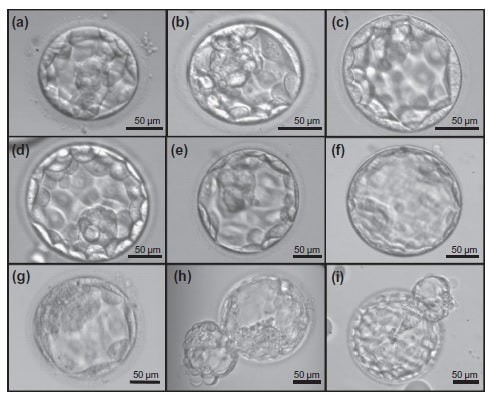 Figure 2. Examples of blastocyst grading: (a) 3AA blastocyst; (b) 3AB blastocyst; (c) 3BA blastocyst; (d) 4AA blastocyst; (e) 4AB blastocyst; (f) 4BA blastocyst; (g) 4CC blastocyst; (h) 5AA blastocyst; (i) 5CA blastocyst. For details of the EH stages and ICM and TE grades, see Materials and methods from Van den Abbeel (11). BLASTOCYST ASSESSMENT BY TIME-LAPSE TECHNOLOGY As previously explained, the most accepted blastocyst grading system is Gardner’s (17), based on the degree of blastocyst expansion and the morphological appearance of both the ICM and TE. However, since embryo development is a dynamic process, conventional grading practices may not detect subtle differences in morphology, which changes significantly over a time span of only a few hours (18). In order to obtain a complete picture of morpho-kinetic events occurring during embryo development a time-lapse system is needed. This technology offers continuous monitoring of embryos rather than just a limited number of discrete observations annotated through conventional assessment. Besides, time-lapse allows embryos to be cultured uninterruptedly, thus getting rid of embryo trafficking from and into the incubator (19). Nevertheless, the actual new and unique contribution of morpho-kinetics is the ability to predict how likely is for a zygote to reach the blastocyst stage in vitro. Several algorithms based on parameters detected by time-lapse, such as early divisions of cleavage-stage embryo, have recently been developed in IVF laboratories to predict blastocyst formation (20). In addition, some authors have made an effort to take time-lapse usefulness further, for instance, to predict the ploidy status of pre-implantation embryos (21, 22). Implantation potential of blastocysts can be evaluated by means of time-lapse during its development. In this regard, three main events are currently being investigated: duration of both compaction and blastulation plus number of blastocyst collapse events (19, 23, 24). Duration of compaction After several cell divisions during the initial stages of embryonic development, the intercellular boundaries become obscured in a process called compaction, which maximizes the intercellular contact and gives rise to the morula (25). Although the compaction of embryos has not received sufficient attention in the IVF field, some studies have focused on the relationships between compaction patterns and embryo developmental potential. Embryos that begin to compact before the eight-cell stage exhibit aberrant in vitro development. Conversely, embryos that complete compaction on day 5 have a lower ability to develop into high-quality blastocysts than those that compact on day 4 (26). These results suggest that the compaction patterns of embryos can facilitate the prediction of their ability to develop both in vitro and in vivo. An interesting work on this issue has been recently published by Mizobe and collaborators (23). The study retrospectively examined the outcome of 299 embryos from 243 patients, which were transferred at blastocyst stage. The whole early development was analysed by comparing morpho-kinetic parameters between implanted and non-implanted embryos, and measuring the time length of specific events, particularly of embryo compaction. Compaction length was calculated by using values of beginning and end of compaction. Beginning of compaction was considered as the time point when the intercellular boundaries became diffuse somewhere in the embryo, while fully compaction was defined as the point when blastomeres were finally unified into one cluster. Compaction length was significantly shorter in blastocysts resulting in pregnancies compared to those that failed to do so. These results indicate a correlation between the length of compaction and implantation potential. This finding is in agreement with the results from previous studies, which observed that the compaction patterns of embryos affected the rates of good-quality blastocyst formation and implantation (26, 27, 28). By contrast, some studies have reported that compaction time of embryos does not affect clinical pregnancy rates (29, 30). 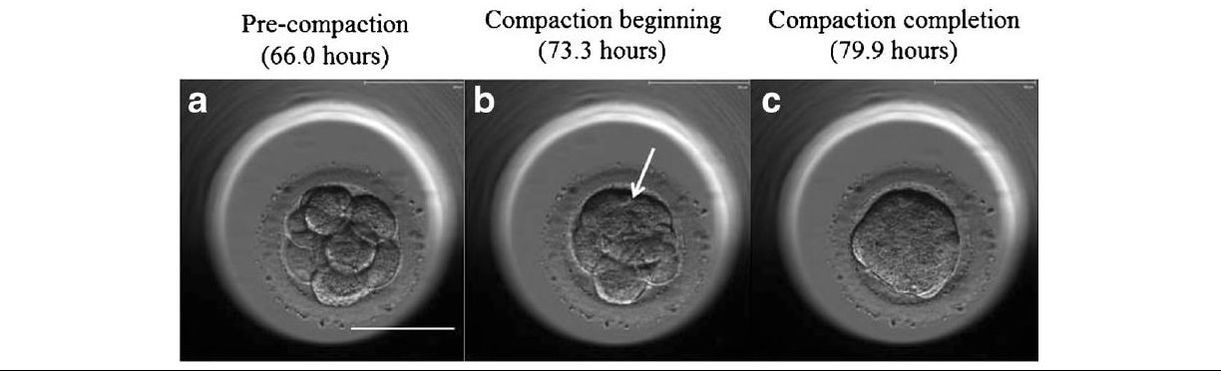 Figure 3. Optimal compaction timing of a blastocyst according to Mizobe and colleagues (23). Duration of blastulation Blastulation is the process through which a morula becomes a blastocyst. Two different structures will arise to form the blastocyst out of the compacted blastomeres of the morula. The first sign of blastulation is compaction and differentiation of the outer blastomeres, forming the TE. This compaction gives the structure a watertight condition, allowing the fluid later secreted to be contained (31). Then, a different group of blastomeres normally located at the centre of the morula start to get closely attached to each other by the formation of Gap junctions, thus facilitating cell communication. It is these cells that differentiate into the ICM (the future embryoblast) and acquire a polarized location at one edge of the embryo. Such polarization creates a cavity, the blastocoel, and gives rise to the structure termed blastocyst. The trophoblasts (TE cells), in turn, continuously pump fluid into the blastocoel, which results in an enhanced size of the blastocyst. This increased volume leads the embryo to hatch through the zona pellucida (32). A recent study conducted by Mumusoglu analysed whether time-lapse morpho-kinetic variables differ among those euploid blastocysts that result in ongoing pregnancy after single embryo transfer (24). For that purpose, 129 patients who had been transferred a single embryo after an ICSI cycle with PGS were considered. Embryos were cultured in a time-lapse incubator up to the moment of TE biopsy, and 23 time-lapse morpho-kinetic parameters were annotated. After biopsy, blastocysts were vitrified and transferred within the next cycle. When comparing all time-lapse parameters, only blastulation time was statistically different: it had lasted shorter in successfully implanted blastocysts than in those that had not implanted. Blastulation time was calculated as the interval from initiation of blastulation up to full blastocyst formation (33, 34). Even though only a few studies have genetically tested euploid blastocysts (21, 22), all of them have pointed out that faster-developing euploid blastocysts might exhibit higher implantation potential. Even so, further large-scale studies are needed in order to confirm such an association (24). Blastocyst collapse events The phenomenon of blastocyst collapse is actually the shrinkage caused by the efflux of the blastocoel fluid due to the loss of cell bindings along the TE. When blastocysts expand, fluid gradually accumulates in the blastocoel -mediated by the sodium pump (Na+/K+-ATPase) (35), resulting in an increased pressure on both the TE and the zona. In parallel, TE cells produce lysins that are involved in the zona weakening and hatching. Formerly to implantation, the embryo needs to leave the zona behind, place adjacent to the endometrial epithelium and then make first contact with the uterus (36). Thus, embryo hatching from the zona is thought to be related to collapse-expansion cycles. By using a time-lapse monitoring system, it has been observed that many of the human blastocysts that reach stage 5 of expansion experience one or more collapse events of the blastocoel cavity, producing a separation of part (if not all) of the TE cells from the zona (19). In a study conducted by IVI Valencia and IVI Murcia clinics (19), blastocyst collapse was analysed to determine its potential influence on reproductive outcomes and whether it may serve for prognostic purposes. 460 patients and data from over 500 blastocysts known to have implanted were included in the study. Blastocyst collapse was considered to have occurred if the separation between TE and the zona pellucida was higher than 50% of the volume. Blastocysts that had experienced just one collapse event were found to present a significantly reduced implantation potential when compared to those transferred after having experienced none. The authors proposed that the molecular mechanisms underlying this association could be related to the mechanical stress suffered from by the embryo, which could result in an excessive energy consumption that would adversely affect the consequent development (19). 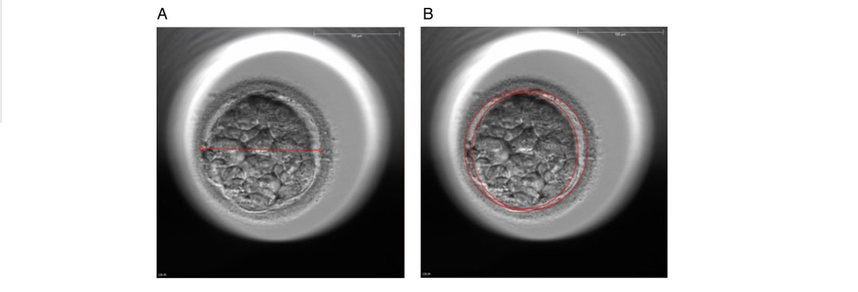 Figure 4. Drawing tools used with Embryovieverw for blastocyst collapse evaluation. First, a line was drawn across the embryo diameter (A). Then, the two circumferences that define the contracted blastocyst and the inner surface of the zona pellucida were outlined (B) [for more details, go to Materials and methods from Marcos (19)]. In spite of the data discussed above, the negative association between blastocyst collapse and implantation potential is not yet clear. In a report by Bodri and colleagues (37), blastocysts were classified according to the number of collapses: embryos with no collapses represented 54% of the total, 22% of the embryos had experienced one single collapse, and multiple collapse events occurred in 24% of the blastocysts. Whereas the live birth rate was observed to decrease as the number of embryo collapse increased, multivariate analyses suggested blastocyst collapse not to be a significant predictor. Rather, it was found to be a confounding factor, along with other morpho-kinetic variables such as time up to two-cell division completion and female age. Therefore, it was concluded that blastocyst collapse patterns should not be evaluated alone without stronger predictors of reproductive outcomes being taken into account (37). TIME-LAPSE AS A MEANS TO EVALUATE EMBRYO QUALITY As previously exposed, the use of time-lapse technology is recently common in embryology laboratories because of its noticeable potential for enhancing embryo selection. Using these technologies, Desai et al. analysed possible kinetic differences between embryos with limited potential and those that accomplished in vitro blastocyst formation and/or implantation (38). Certain parameters such as time of pronuclear formation and cleavage stage were found to be different in embryos reaching blastocyst stage vs. poor-quality embryos. Moreover, a large number of embryos were found to present multinucleation and reverse cleavage, but they were able to form a blastocyst with optimal criteria for freezing (38), which resembles previous reports on the dynamic nuclear formation of blastocysts by Ergin and coauthors (39). With respect to blastocyst formation, Motato et al. (2016) proposed two models to classify embryos based on their probability of reaching blastocyst stage and implantation (40). However, the study was limited by parameters such as subjective criteria from different clinics with different culture media (40). Consequently, it would be reasonable to keep on research on this subject in order to achieve a consensus regarding embryo classification and implantation potential (40). CONSIDERING DAY-2 AND DAY-3 EMBRYO MORPHOLOGY BEFORE DAY-5 TRANSFER Even though blastocyst stage is currently widely accepted as the optimal moment for embryo transfer, cleavage stage has been traditionally regarded as the right moment in global practice. In fact, it still continues to be so in some laboratories, and early transfer into the uterus has been proposed to be advantageous to the embryo due to the limited time exposed to the in vitro environment (41). However, there exist two main arguments supported by extensive scientific literature to explain why blastocyst transfer after extended culture has advantages over the traditional cleavage-stage transfer: First of all, when the embryo arrives to the uterus in natural conditions it has already reached morula stage, which corresponds to, at least, day 4 of in vitro culture (42). This means blastocyst stage is the most physiologically compatible stage for transfer, since it allows a better synchronization between embryonic stage and endometrial receptivity (43) [you can read more about the optimal day for embryo transfer in our previous article here]. Secondly, several studies have reported higher implantation potential for blastocysts compared to cleavage-stage embryos (6, 41), the first transferred blastocyst being reported in 1995 (44). Furthermore, some authors have postulated that a large proportion of morphologically normal day-3 embryos are actually chromosomally abnormal or mosaic, which may contribute to the 80-90% rate of implantation failure observed after cleavage-stage embryo transfer (45). EXTENDING EMBRYO CULTURE UP TO BLASTOCYST STAGE Considering the need for further studies on the subject, and the fact that day-3 embryos can actually implant and develop successfully, does it really make sense to extend embryo culture up to blastocyst stage? As above-stated, morphologically normal embryos may actually present chromosome abnormalities, which proves the insufficiency of morphological criteria to evaluate implantation rates (46). Because of embryo plasticity, the proportion of chromosomally abnormal cells varies within the culture; corrupted cells can be eliminated, thus resulting in a good-quality blastocyst developing from a poor-quality cleavage-stage embryo (47). Some studies have evaluated pregnancy rates derived from transfers of blastocysts with previous poor quality as cleavage-stage embryos, finding an approximate success rate of 45% after culture and freezing of embryos at an early stage for another cycle. The conclusion of this being a valid practice to avoid the repetition of IVF-ICSI treatments (48) agrees with recent findings showing that low-scoring day-2 or 3 embryos, which are not considered transferable, can still result in successful blastulation and end up in a live birth (49, 50). All this said, the right question now would be: should day-2 and day-3 embryo morphology be considered before transfer at day 5 when blastocysts reach a similar good quality? A recent retrospective study by Herbemont has suggested that only the quality of the transferred blastocyst may be predictive of the subsequent clinical outcome, whereas morphological aspects at day 2 or day 3 have limited interest (51). These same results had previously been observed by Guerif; even though early morphological parameters were relatively helpful to predict blastocyst development, their value to predict blastocyst morphology was limited, and so they provided no significant additional information that could prognosticate blastocyst implantation and live birth rates (6). A few years earlier and with the same goal in mind, Zech and coauthors carried out a prospective randomized study in which they compared ongoing pregnancy rates per single embryo transfer between day 3 and day 5. When good-quality embryos were available, pregnancy rates were found to be higher after blastocyst transfer. Therefore, the authors concluded that morphological criteria-based seleccion at day 3 may not be a suitable procedure when just one embryo is to be transferred out of a cohort of all morphologically good ones. Thus, and as stated by the authors, extending embryo culture up to day 5 may result in a better strategy in order to correctly identify and select those embryos with higher implantation potential, provided there is a sufficient number of top-quality eight-cell embryos available (52). On the contrary, a study performed by Silber (2014) found that blastocysts arising from poor-quality embryos displayed lower implantation and pregnancy rates compared to good-quality embryos. These discrepancies could be due to different criteria used to score embryo quality (53). So, in order to minimise discrepancies between studies, the use of time-lapse is currently established as a common approach to evaluate embryo morpho-kinetics. In fact, reduction in the time of embryo exposure to the environment outside the incubator has been demonstrated to enhance both embryo quality and blastulation rates (54). Nevertheless, to answer the question previously postulated, a prospective randomized study would be needed that compare at least two similar good-quality blastocysts, one arising from a good-quality day-2/3 embryo, and to the other from a poor-quality one (51). CONCLUSIONS It is important to take into account that the main population features of different patients, such as paternal age, maternal BMI, parental smoking or cause of infertility may influence clinical outcomes. Moreover, certain methodological aspects also need to be considered, like blastocyst evaluation by the same personal (in order to minimize variation) or the consistent use of the same type of culture media (to avoid potential effects on birth-weight and other traits), just as previously suggested (4). A universal embryo grading system needs to be validated, before widespread implementation in IVF laboratories. Also, it has not yet been clearly established which morphological feature of blastocysts (expansion, TE or ICM) is the most reliable as a predictive factor for post-transfer implantation success. Consequently, there is still a debate between authors about the true outcomes of single transfer of low-quality blastocysts (4). Morpho-kinetics assessment, along with chromosomal screening, may ultimately help identify euploid embryos with the highest developmental potential (55). Since these features are susceptible of being affected by in vitro culture conditions, each embryology laboratory should define their own cut-off points in order to standardise time-lapse variables (24). Finally, it should be taken into account the fact that embryo quality is not the only parameter with influence on implantation rates; endometrial receptivity is also involved, and it may be greatly determined by a variety of factors (56) [to learn more about endometrium status and receptivity, read our previous post here]. REFERENCES
1- Souza Setti A, Ferreira RC, Paes de Almeida, Ferreira Braga D, de Cassia Savio Figueira R, Iaconelli A Jr, Borges E Jr. Intracytoplasmic sperm injection outcome versus intracytoplasmic morphologically selected sperm injection outcome: a meta-analysis. Reprod Biomed Online 2010;21:450-455. 2- Meldrum DR. Blastocyst transfer: a natural evolution. Fertil Steril 1999;72:216–7. 3- Balaban B, Urman B, Sertac A, Alatas C, Aksoy S, Mercan R. Blastocyst quality affects the success of blastocyst-stage embryo transfer. Fertil Steril 2000;74:28-287. 4- Bouillon C, Celton N, Kassem S, Frapsauce C, Guérif F. Obstetric and perinatal outcomes of singletons after single blastocyst transfer: is there any difference according to blastocyst morphology? Reprod Biomed Online 2017;35:197–207. [Internet]. 5- Stecher A, Vanderzwalmen P, Zintz M, Wirleitner B, Schuff M, Spitzer D, Zech NH. Transfer of blastocysts with deviant morphological and morphokinetic parameters at early stages of in-vitro development: a case series. Reprod Biomed Online 2014;28:424-435. 6- Guerif F, Lemseffer M, Leger J, Bidault R, Cadoret V, Chavez C, Gasnier O, Saussereau MH, Royere D. Does early morphology provide additional selection power to blastocyst selection for transfer? Reprod Biomed Online 2010;21:510– 519. 7- Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology. The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum Reprod 2011;26:1270 –1283. 8- Gardner DK, Schoolcraft WB, Wagley L, Schlenker T, Stevens J, Hesla J. A prospective randomized trial of blastocyst culture and transfer in in-vitro fertilization. Hum Reprod 1998;13:3434-3440. 9- Balaban B, Yakin K, Urman B. Randomized comparison of two different blastocyst grading systems. Fertil Steril 2006;85:559– 563. 10- Richardson A, Brearley S, Ahitan S, Chamberlain S, Davey T, Zujovic L, Hopkisson J, Campbell B, Raine-Fenning N. A clinically useful simplified blastocyst grading system. Reprod Biomed Online 2015;31:523–530. 11- Van den Abbeel E, Balaban B, Ziebe S, Lundin K, Cuesta MJ, Klein BM, et al. Association between blastocyst morphology and outcome of singleblastocyst transfer. Reprod Biomed Online 2013;27:353–61. 12- Shapiro BS, Daneshmand ST, Garner FC, Aguirre M, Thomas S. Large blastocyst diameter, early blastulation, and low preovulatory serum progesterone are dominant predictors of clinical pregnancy in fresh autologous cycles. Fertil Steril 2008;90:302–9. 13- Shapiro BS, Richter KS, Harris DC, Daneshmand ST. A comparison of day 5 and 6 blastocyst transfers. Fertil Steril 2001;75:1126–30. 14- Ahlström A, Westin C, Reismer E, Wikland M, Hardarson T. Trophectoderm morphology: an important parameter for predicting live birth after single blastocyst transfer. Hum Reprod 2011;26:3289– 3296. 15- Hill MJ, Richter KS, Heitmann RJ, Graham JR, Tucker MJ, Decherney AH, Browne PE, Levens ED. Trophectoderm grade predicts outcomes of single-blastocyst transfers. Fertil Steril 2013;99: 1283–1289. 16- Almagor M, Harir Y, Fieldust S, Or Y, Shoham Z. Ratio between inner cell mass diameter and blastocyst diameter is correlated with successful pregnancy outcomes of single blastocyst transfers. Fertil Steril 2016;106: 1386–1391. [Internet] 17- Gardner DK, Lane M, Stevens J, Schlenker T, Schoolcraft WB. Blastocyst score affects implantation and pregnancy outcome: towards a single blastocyst transfer. Fertil Steril. 2000;73(6):1155–8. 18- Cruz M, Garrido N, Herrero J, Pérez-Cano I, Muñoz M, Meseguer M. Timing of cell division in human cleavage-stage embryos is linked with blastocyst formation and quality. Reprod Biomed Online. 2012;25(4):371–81. 19- Marcos J, Pérez-Albalá S, Mifsud A, Molla M, Landeras J, Meseguer M. Collapse of blastocysts is strongly related to lower implantation success: a time-lapse study. Hum Reprod. 2015;30(11):2501–8. 20- Meseguer M, Herrero J, Tejera A, Hilligsøe KM, Ramsing NB, Remohí J. The use of morphokinetics as a predictor of embryo implantation. Hum Reprod. 2011;26(10):2658–71. 21- Yang Z, Zhang J, Salem SA, Liu X, Kuang Y, Salem RD, et al. Selection of competent blastocysts for transfer by combining time-lapse monitoring and array CGH testing for patients undergoing preimplantation genetic screening: a prospective study with sibling oocytes. BMC Med Genomics. 2014;7(1):38. 22- Kofinas JD, Tiegs A, Kramer YG, McCulloh DH, Grifo JA. Do Time Lapse Morphokinetic (TLM) Parameters Distinguish between Good Versus Poor Prognosis Embryos of Known Ploidy Status? Fertil Steril. 2015;103(2):e27. 23- Mizobe Y, Ezono Y, Tokunaga M, Oya N, Iwakiri R. Selection of human blastocysts with a high implantation potential based on timely compaction. J Assist Reprod Genet. 2017;34(8):991-997. 24-Mumusoglu S, Ozbek IY, Sokmensuer LK, Polat M, Bozdag G, Papanikolaou E, et al. Duration of blastulation may be associated with ongoing pregnancy rate in single euploid blastocyst transfer cycles. Reprod Biomed Online. 2017;35(6):633-639. 25- Fleming TP, Sheth B, Fesenko I. Cell adhesion in the preimplantation mammalian embryo and its role in trophectoderm differentiation and blastocyst morphogenesis. Front Biosci. 2001;6(1):D1000–7. 26- Iwata K, Yumoto K, Sugishima M, Mizoguchi C, Kai Y, Iba Y, et al. Analysis of compaction initiation in human embryos by using time-lapse cinematography. J Assist Reprod Genet. 2014;31(4):421–6. 27- Skiadas CC, Jackson K V, Racowsky C. Early compaction on day 3 may be associated with increased implantation potential. Fertil Steril. 2006;86(5):1386–91. 28- Tao J, Tamis R, Fink K, Williams B, Nelson-White T, Craig R. The neglected morula/compact stage embryo transfer. Hum Reprod. 2002;17(6):1513–8. 29- Chamayou S, Patrizio P, Storaci G, Tomaselli V, Alecci C, Ragolia C, et al. The use of morphokinetic parameters to select all embryos with full capacity to implant. J Assist Reprod Genet. 2013;30(5):703–10. 30-Kirkegaard K, Kesmodel US, Hindkjær JJ, Ingerslev HJ. Time-lapse parameters as predictors of blastocyst development and pregnancy outcome in embryos from good prognosis patients: a prospective cohort study. Hum Reprod. 2013;28(10):2643–51. 31- Forgacs G, Newman SA. Biological physics of the developing embryo. Cambridge University Press; 2005. 32- Larsen WJ. Human embryology. Churchill Livingstone; 2001. 33- Campbell A, Fishel S, Bowman N, Duffy S, Sedler M, Hickman CFL. Modelling a risk classification of aneuploidy in human embryos using non-invasive morphokinetics. Reprod Biomed Online. 2013;26(5):477–85. 34- Ciray HN, Campbell A, Agerholm IE, Aguilar J, Chamayou S, Esbert M, et al. Proposed guidelines on the nomenclature and annotation of dynamic human embryo monitoring by a time-lapse user group. Hum Reprod. 2014;29(12):2650–60. 35- Baltz JM, Smith SS, Biggers JD, Lechene C. Intracellular ion concentrations and their maintenance by Na+/K+-ATPase in preimplantation mouse embryos. Zygote. 1997;5(1):1–9. 36- Biggers JD. Reflections on the culture of the preimplantation embryo. Int J Dev Biol. 2004;42(7):879–84. 37- Bodri D, Sugimoto T, Serna JY, Kawachiya S, Kato R, Matsumoto T. Blastocyst collapse is not an independent predictor of reduced live birth: a time-lapse study. Fertil Steril. 2016;105(6):1476–83. 38-Desai N, Ploskonka S, Goodman LR, Austin C, Goldberg J, Falcone T. Analysis of embryo morphokinetics, multinucleation and cleavage anomalies using continuous time-lapse monitoring in blastocyst transfer cycles. Reprod Biol Endocrinol. 2014;12:54. 39- Ergin EG, Calişkan E, Yalçinkaya E, Oztel Z, Cökelez K, Ozay A, et al. Frequency of embryo multinucleation detected by time-lapse system and its impact on pregnancy outcome. Fertil Steril. 2014;102(4):1029-33.e1. 40- Motato Y, de los Santos MJ, Escriba MJ, Ruiz BA, Remohí J, Meseguer M. Morphokinetic analysis and embryonic prediction for blastocyst formation through an integrated time-lapse system. Fertil Steril. 2016;105(2):376-84.e9. 41- Glujovsky D, Farquhar C, Quinteiro Retamar AM, Alvarez Sedo CR, Blake D. Cleavage stage versus blastocyst stage embryo transfer in assisted reproductive technology. Cochrane Database Syst Rev. 2016(6):CD002118. 42- Croxatto HB, Fuentealba B, Díaz S, Pastene L, Tatum HJ. A simple nonsurgical technique to obtain unimplanted eggs from human uteri. Am J Obstet Gynecol. 1972;112(5):662-8. 43- Valbuena D, Martin J, de Pablo JL, Remohí J, Pellicer A, Simón C. Increasing levels of estradiol are deleterious to embryonic implantation because they directly affect the embryo. Fertil Steril. 2001;76(5):962-8. 44- Edwards RG, Brody SA. History and ethics of assisted human conception. Principles and Practice of Assisted Human Reproduction. Philadelphia: WB Sauders, 1995:17–47. 45- Magli MC, Gianaroli L, Munné S, Ferraretti AP. Incidence of chromosomal abnormalities from a morphologically normal cohort of embryos in poor-prognosis patients. J Assist Reprod Genet. 1998;15(5):297-301. 46- Meseguer M, Rubio I, Cruz M, Basile N, Marcos J, Requena A. Embryo incubation and selection in a time-lapse monitoring system improves pregnancy outcome compared with a standard incubator: a retrospective cohort study. Fertil Steril. 2012;98(6):1481-9.e10. 47- Fragouli E, Alfarawati S, Spath K, Jaroudi S, Sarasa J, Enciso M, et al. The origin and impact of embryonic aneuploidy. Hum Genet. 2013;132(9):1001-13. 48- Kaartinen N, Das P, Kananen K, Huhtala H, Tinkanen H. Can repeated IVF-ICSI-cycles be avoided by using blastocysts developing from poor-quality cleavage stage embryos? Reprod Biomed Online. 2015;30(3):241-7. 49- Hershko Klement A, Ovadia M, Wiser A, Berkovitz A, Shavit T, Nemerovsky L, et al. What we learned from extended culture of 'rejected' day-3 cleavage stage embryos: a prospective cohort study. J Ovarian Res. 2017;10(1):35. 50- Sallem A, Santulli P, Barraud-Lange V, Le Foll N, Ferreux L, Maignien C, et al. Extended culture of poor-quality supernumerary embryos improves ART outcomes. J Assist Reprod Genet. 2017. 51- Herbemont C, Sarandi S, Boujenah J, Cedrin-Durnerin I, Sermondade N, Vivot A, et al. Should we consider day-2 and day-3 embryo morphology before day-5 transfer when blastocysts reach a similar good quality? Reprod Biomed Online. 2017;35(5):521-8. 52- Zech NH, Lejeune B, Puissant F, Vanderzwalmen S, Zech H, Vanderzwalmen P. Prospective evaluation of the optimal time for selecting a single embryo for transfer: day 3 versus day 5. Fertil Steril. 2007;88(1):244-6. 53- Silber, S., 2014. Blastocysts that came from better quality day 3 embryos give higher pregnancy rates than blastocysts that derive from poor quality day 3 embryos. Ovarian Club, Paris. 54- Zhang JQ, Li XL, Peng Y, Guo X, Heng BC, Tong GQ. Reduction in exposure of human embryos outside the incubator enhances embryo quality and blastulation rate. Reprod Biomed Online. 2010;20(4):510-5. 55- Zaninovic N, Irani M, Meseguer M. Assessment of embryo morphology and developmental dynamics by time-lapse microscopy: is there a relation to implantation and ploidy? Fertil Steril. 2017;108(5):722-9. 56- Valbuena D, Valdes CT, Simon C. Introduction: Endometrial function: facts, urban legends, and an eye to the future. Fertil Steril. 2017;108(1):4-8. "The underlying cause for endometriosis is likely to present a multifactorial origin" Authors: Javier del Río, Noemi Díaz & Edel Rocher 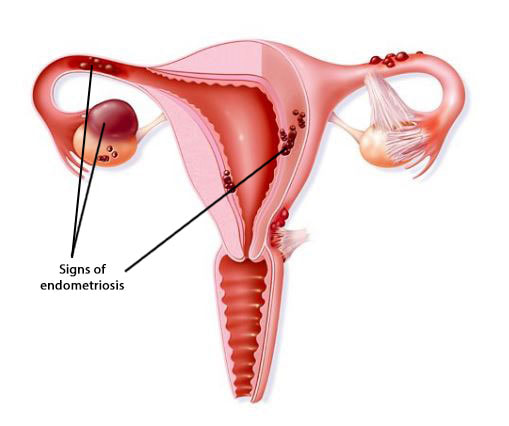 Figure 1. Illustration showing evidences of endometriosis found in specific locations of the female reproductive organs [1]. Endometriosis is defined as the presence of endometrial tissue outside the uterus, which induces a chronic inflammatory reaction. It is generally assumed that lesions arise through retrograde endometrial tissue loss during menstruation, coelomic metaplasia and lymphatic spread in immunologically and genetically susceptible individuals. Associated symptoms usually have an impact on the general physical, mental and social well-being (2,3,4). This condition is found in women from all ethnic and social groups, predominantly in those of reproductive age, which may represent an estimation of up to 176 million women all over the world. The prevalence of endometriotic disease seems to be around 5%, normally reaching a peak somewhere between ages 25 and 35. Among women aged 15-49, an annual incidence of endometriosis of 0.1% has been reported (2,3,4). Which symptoms may be indicative of the disease? Women who suffer from endometriosis typically experiment abdominal pain and related symptoms such as dysmenorrhoea, dyspareunia, heavy menstrual bleeding, non-menstrual pelvic pain, painful ovulation, dyschezia and dysuria, as well as chronic fatigue. Endometriotic lesions are followed by denervation and reinnervation, which represent accompanying changes in the central nervous system (central sensitization), thus generating a chronic pain syndrome. The extent of the disease varies from a few, small lesions on otherwise normal pelvic organs to large, ovarian endometriotic cysts (endometriomas) and/or extensive fibrosis and adhesion formation, causing marked distortion of the pelvic anatomy (2,3). As a consequence, establishing the diagnosis of endometriosis on the basis of symptoms alone may result difficult, not only due to their high degree of variation, but also because there is so much overlapping with other conditions such as irritable bowel syndrome or pelvic inflammatory disease. Therefore, it is common to come across a delay of several years between the onset of symptoms and a definitive diagnosis. In this respect, the revised scoring system of the American Society for Reproductive Medicine is normally employed to determine the disease stage (ranging from I, indicating minimal disease, to IV, indicating severe disease). This system is based on the type, location and appearance of the lesions, as well as the depth of the invasion caused and the extent of disease and adhesions (2,5,6). However, endometriosis symptoms do not always correspond to observations from laparoscopic exploration. The severity of these symptoms increase with age, along with the probability of a proper diagnosis. Moreover, the incidence of endometriosis peaks in women in their 40s (7). Which are the possible causes of the disease? Even though the underlying cause for endometriosis remains uncertain, it is likely to present a multifactorial origin. This includes inflammation enhanced by genetic causes, which may be epigenetically regulated, as well as triggered by exposure to environmental factors (3). So, it seems evident why pathophysiology of endometriosis keeps being a subject of debate. It remains unclear whether endometrial and endometriotic cells are genetically different, or whether such differences are the consequence of distinct environments or due to preexisting immunological defects (8). However, several hypotheses have been proposed to explain the origin of the disease:
This was the first hypothesis proposed, put forth as the main cause for the pathogenesis of endometriosis. Retrograde menstruation appears when viable endometrial fragments are driven through the Fallopian tubes, possibly by a pressure gradient originating from dys-synergic uterine contractions. Once these fragments reach the peritoneal cavity they can implant, grow and invade pelvic structures. The likelihood of this event is influenced epidemiologically by any menstrual, reproductive or personal factor that may augment pelvic contamination by regurgitated endometrium (Figure 1). Further support for this aetiology is derived from studies of obstructed or compromised outflow tracts (9,10,11). However, this event occurs in 76–90% of women with patent Fallopian tubes, and not all of them suffer from endometriosis (11).
The monthly regeneration of the endometrium after menstrual shedding and re-ephitelialization after parturition or surgical curettage supports the existence of a stem cell pool. The involvement of these cells in the formation of endometriotic deposits could be the result of abnormal translocation of the normal stratum basalis of the endometrium via retrograde menstruation. Some group of cells may deposit and survive in the peritoneal cavity after retrograde flow, and may reactivate during adolescence in response to ovarian hormones. However, there are currently no data on the amount of endometrial stem/progenitor cells in neonatal period compared to the adult endometrium (11).
This theory postulates that endometriosis originates from the metaplasia of specialised cells present in the mesothelial lining of the visceral and abdominal peritoneum. Hormonal or immunological factors are thought to stimulate the transformation of normal peritoneal tissue/cells into endometrium-like tissue. The theory may explain the occurrence of endometriosis in pre-pubertal girls. However, the usual driving force for endometrial growth, oestrogen, is not present in pre-pubertal girls, and therefore this condition may be different from the endometriosis found in women of reproductive age. Nevertheless, this theory is imperfect due to endometriotic lesions being found in areas outside the course of the Müllerian duct (4,11).
Genetic factors represent about half of the variation in endometriosis risk, with an estimate of heritability of 51%. Meta-analyses of the few genome-wide studies performed in the past few years have provided evidence of a robust association of endometriosis with seven risk loci. Among them, WNT4, CDKN2B-AS1 and GREB1 are of particular interest due to their gene-based ranking, known pathophysiology and proximity to SNPs of genome-wide significance. Therefore, these genes represent strong candidates for further studies on endometriosis (4,12,13). Inflammation is an additional typical feature of endometriosis, since the presence of ectopic tissue in the peritoneal cavity is associated with overproduction of prostaglandins, cytokines and chemokines. Macrophages infiltrating the ectopic lesions express typical markers of alternative activation, favouring the growth of the lesions and promoting associated angiogenesis. Overproduction of reactive oxygen species (ROS) is also accompanied by a decreased level of antioxidants that usually eliminates these molecules. Thus, the resulting accumulation of ROS may contribute to the propagation and maintenance of endometriosis and associated symptoms (4,10,11). Oestrogens fuel ectopic endometrium growth, and alterations in oestrogen signalling have been associated with the disease. In fact, it is known that oestradiol produced both in the steroidogenic organs and locally in the endometriotic implants through aromatase expression promote the growth of endometrial tissue ectopically (4,11). This ectopic tissue has been consistently shown to express different levels of oestrogen receptors (ER) α and β than eutopic tissue, being ERβ highly present in ectopic tissue (4). Deficient methylation of the promoter of the ERβ-encoding gene has been suggested to result in pathological overexpression of ERβ in endometriosis, which in turn suppresses ERα expression and diminishes oestradiol-mediated induction of the progesterone receptor in endometriotic cells. This mechanism is thought to contribute to resistance to selective actions of progesterone in these cells, which is manifested by perturbations in a number of downstream progesterone target genes (4,11). Progesterone normally triggers a uterine endometrial response characterized by inhibition of oestrogen-dependent proliferation of epithelial cells, secretory maturation of the glands, and transformation of stromal cells into specialized decidual cells. Moreover, progesterone transiently induces a receptive phenotype in endometrial epithelial cells essential for embryo implantation [you can read more about the role of these hormones in the endometrium in our previous publication here]. As a consequence of progesterone resistance, genes critical to these events, such as prolactin for decidual response or glycodelin for embryo implantation, are dysregulated in the endometrium of affected women. Endometriosis-derived inflammation, in turn, could induce progesterone resistance by altering the signalling pathway of this hormone through mechanisms of competition or interference with proinflammatory transcriptional factors (4,11,14,15,16). The observation that autoimmune diseases tend to be more common in women with endometriosis supports the hypothesis that pathogenesis of this condition may involve a defective immune response in these patients. Women suffering from endometriosis present higher concentration of activated macrophages, decreased cellular immunity and repressed NK cell function (11). Endometriosis and infertility The relationship between endometriosis and infertility has been debated for many years. Infertile women are 6-8 times more likely to suffer from endometriosis than fertile women (7). Despite extensive research no agreement has been reached, and several mechanisms have been proposed to explain the association between endometriosis and infertility. These mechanisms include distorted pelvic anatomy, endocrine and ovulatory abnormalities, altered peritoneal function and altered hormonal and cell-mediated functions in the endometrium. Major pelvic adhesions or peritubal adhesions disturbing the tube-ovarian liaison and tube patency can impair the release of oocytes from the ovary, inhibit oocyte pickup, or impede the transport of the ovum itself (7). Women with endometriosis may exhibit endocrine and ovulatory disorders, including luteinized unruptured follicle (LUF) syndrome, impaired folliculogenesis, luteal phase defect, and premature or multiple LH surges. Moreover, a complex network of humoral and cellular immunity factors modulates the growth and inflammatory behaviour of ectopic endometrial implants, and so it may have adverse effects on the function of the oocyte, sperm, embryo, or Fallopian tubes (7,16). Management of endometriosis by assisted reproductive technologies There is currently no clear answer to the question of whether endometriosis has a negative impact on IVF outcomes. Several studies have previously suggested poorer outcomes in comparison to control cases, whereas other have shown no significant differences (17). Clinical outcomes for different stages of endometriosis Patients with endometriosis can present different levels of severity, which have a strong influence in the outcomes. Thus, it would be common to find a case of disease at a more severe phase with worse results than a different one presenting less extensive endometriosis (18,19). In stage III/IV endometriosis the prognosis for IVF/ICSI treatments is lower compared to milder stages I/II. (19). In their retrospective study, Opøien et al. highlighted the fact that patients with a more severe form of the disease had fewer oocytes retrieved, despite the higher gonadotropin doses these women had been administered (20). This may be due to endometriosis affecting both oestrogen and progesterone mRNA receptors in granulosa cells (21). Likewise, patients suffering from stage III/IV endometriosis have been reported to show a decrease in implantation and clinical pregnancy rates (22). Endocrinology of endometriosis Measurements of circulating hormone concentrations have shown statistically higher levels of hMG/FSH in endometriosis patients than in control patients. However, both high-quality embryo and clinical pregnancy rates were found to be lower in such patients (23). Similarly, the high expression of PR-A and ER-α in patients with endometriosis might be a leading cause of ovarian dysfunction due to this condition. This means that stages III-IV patients required higher gonadotropin stimulation doses to prevent ovarian dysfunction (24). Endometriomas Endometriomas represent an additional variable that may also affect ART outcomes. They have been reported to affect 17-44% of patients with endometriosis, who exhibit reduced ovarian response due to the low response to gonadotropin stimulation (25). Such response has been widely discussed by different authors. Conclusions from these reports suggest a reduced number of retrieved oocytes in women with bilateral endometriomas, as well as the rates of top-quality embryos, implantation, clinical pregnancy and live birth, when compared to control groups (26,27,28). Gonadotropin stimulation for IVF in the endometriosis context Patients with stage III/IV endometriosis normally undergo stimulation by higher doses of gonadotropin, which consequently results in higher circulating oestradiol levels (24). D'Hooghe and coworkers performed an analysis of this kind of patients and demonstrated that the cumulative endometriosis recurrence rate (CERR) was lower after ovarian hyperstimulation for IVF than after lower-dose ovarian stimulation for IUI. This suggests that temporary exposure to high oestradiol levels during ovarian hyperstimulation for IVF is not a risk factor for endometriosis recurrence in women subjected to ART (29). Previous reports from Benaglia et al. have shown that after 3-6 months of IVF cycles 77% of patients experimented an improvement of the condition, whereas about 11% reported worsening. Also, it should be noted that endometrioma size also remained stable after IVF cycles, which suggests they are not affected by gonadotropin stimulation (30). IVF outcomes after medical therapy for endometriosis Medical interventions have been demonstrated to have benefit in alleviating, if not eliminating, symptoms associated with endometriosis (31). The first thing to control is the prolonged use of GnRHa prior to initiation of gonadotropin stimulation for ART. The extension of GnRHa use up to about 3 months has been reported to exhibit higher implantation rates and significantly higher clinical pregnancy rates (32). This is due to the fact that endometriosis patients were more likely to present aberrant endometrial expression of β3 integrin, and that a 3-month course of GnRHa allowed to rescue up to 64% of the expression (33). Another approach employed to increase integrin expression is the of danazol as a post-treatment after the ovulatory cycle. This has been reported to result in both similar clinical pregnancy and live birth rates to those from patients with normal integrin expression (34). A different medical therapy is the administration of oral contraceptives in women with endometriosis before the initiation of the actual ART treatment. This approach seems to improve clinical outcomes, keeping them comparable to women of similar age without endometriosis. In contrast, ART outcomes are markedly compromised in endometriosis patients who are not pretreated with this method. It is worth to mention that ovarian responsiveness to stimulation was not altered by 6-8 weeks use of oral contraceptives prior to initiation of ART treatment, including poor responders with endometriomas (35). So far, it has not been possible to establish the optimal duration of medical therapy for endometriosis by means of comparative trials. As a consequence, ideal patients who could benefit from medical intervention have not yet been depicted. In spite of this, it would be reasonable to think that those patients suffering from more severe stages of the disease and/or with prior implantation failure might be the best candidates (36). Infertility in women affected by endometriosis may be related to alterations in follicles, poor oocyte quality (...), or even decreased endometrial receptivity. Impact of surgical management of endometriosis on IVF outcomes Previous reports have shown that pre-cycle surgical intervention may result beneficial for the final outcome. Opøien et al. demonstrated that patients with stage I/II endometriosis treated with surgical resection presented higher clinical pregnancy rates than those who had only been subjected to diagnostic laparoscopy before IVF/ICSI (37). Similar data were obtained when evaluating results from a group of 825 endometriosis patients; those treated with surgical resection presented overall significantly higher pregnancy and IVF rates when compared to those who had been subjected to IVF alone, and also to those with no treatment whatsoever (38). Implantation and pregnancy rates have also been reported to increase after resection in patients with deeply invasive endometriosis, although this group needed higher gonadotropin doses for stimulation, and yet fewer oocytes were retrieved (39). However, it is important to keep in mind that research is limited by a variety of factors, such as variations in surgical techniques (i.e., ablation vs. resection), degree of removal of the endometriotic tissue or differences in IVF laboratories (40). Interestingly, previous reports indicate that IVF implantation rates are not affected by the time intervals between surgical interventions for resection of endometriosis in the absence of endometriomas (41). On the other hand, certain studies have reported the negative impact of surgical management of endometriomas on IVF treatments, like the inability to access follicles at oocyte retrieval following precycle resection of endometriomas, or the harmful effect on oocytes after exposure to endometrioma fluid. Nevertheless, most authors refuse these arguments (42), and few studies assert the benefits of surgery in endometriomas larger than 3 cm in order to treat painful symptoms or to facilitate access to the ovary (43). Current data from clinical outcomes suggest that, under controlled circumstances, IVF cycles are not compromised by the presence of endometriosis. The exception to this finding is the fact that patients with ovarian endometriomas showed lower response to gonadotropin stimulation (17). Thorough meta-analyses through early studies revealed lower number of oocytes obtained after egg collection, as well as decreasing fertilization, implantation and pregnancy rates in patients affected by endometriosis after ART treatments (18). However, it is important to note that pregnancy rates were extremely low in those years. Effects of endometriosis on ART outcomes Since endometriosis patients have sometimes been studied simultaneously to other groups of patients such as women suffering from Fallopian tube-related infertility, it is not clear whether endometriosis actually affects implantation rates. Barcelo and coauthors’ explanation is the similar percentage of meiotic abnormalities in in vitro matured oocytes between endometriosis patients and the control group after ovarian stimulation (44). An additional factor that might contribute to confusion is the presence of adenomyosis, which is frequently found in endometriosis patients and could have a deleterious impact on the implantation process (45). Altered ovulation and oocyte production, as well as increased inflammatory cells in the peritoneal fluid of endometriomas have been observed in endometriosis patients (46). In fact, infertility in women affected by endometriosis may be related to alterations on follicles, poor oocyte quality and related failures during subsequent embryogenesis, or even decreased endometrial receptivity. Such hypothesis is supported by the altered progesterone and cytokine concentrations found in the follicular fluid from these patients (44). Embryos derived from affected women are likely to develop more slowly, compared to embryos derived from women suffering from tubal disease (45). It is worth to mention that women with moderate to severe endometriosis who receive oocytes from healthy donors seem to present normal endometrial receptivity and pregnancy rates. Conversely, when donor oocytes from endometriosis-affected women are transferred into healthy women, implantation rates are lower and embryo quality is reduced (46). Further studies are needed, though, in order to determine pregnancy rates from donors in different conditions and disease stages (16). Delayed histological maturation or biochemical disturbances may lead to endometrial dysfunction, too. This is concluded from reports that show uterine implantation being affected by changes in receptivity on an endometriosis background (17). Endometrial expression of the adhesion molecule αvβ integrin has been observed to be reduced during the time of implantation in some endometriosis-affected women. Additionally, the synthesis of the endometrial ligand for L-selectin has also been observed to be affected in some patients (16). L-selectin is a trophoblast surface-coating protein, which may explain the altered implantation rates observed in such cases (47). The observation of luteal phase disruption associated to endometriosis may be due to dysregulation of the progesterone receptor, as well as an effect on progesterone target genes that consequently results in a decrease in endometrial receptivity (17,47). Sperm quality and/or function is also reduced, which has been proposed to be due to inflammatory/toxic effects of the peritoneal fluid, along with a higher amount of activated macrophages. Not only are these effects harmful to oocytes and sperm, but toxic to the embryo (17). Treatment of endometriosis-associated infertility IVF is currently the most effective treatment for endometriosis-associated infertility (17). However, comparison of data on the effectiveness of IVF for endometriosis patients vs. patients suffering from other causes of infertility is not without controversy. Reports from the Society of Assisted Reproductive Technology (SART) have shown that the average delivery rate per retrieval for patients undergoing IVF-ET is 39.1% for endometriosis-affected women, as opposed to 33.2% for women with other causes of infertility (48). Pre-treatment ovulation suppression is a possibility to be considered in order to enhance suppression of inflammatory cytokines, as well as to reduce the presence of disease signs prior to any form of ART. Nevertheless, further research will be required on patients with endometriomas to assess their effect on IVF/ICSI, and to elucidate whether pre-ART surgical intervention may increase success rates (48). Potential treatments in the future There is currently no consensus on whether performing surgery prior to undergoing ART is fundamental for achieving pregnancy. Either way, there is a priority in terms of age in endometriosis cases (17). There are, however, some novel medical therapies, such as immunoconjugate (ICON) and aromatase inhibitors. ICON targets aberrantly expressed tissue factor on endometriotic endothelium, causing regression of the established disease (most likely by devascularization), which seems to improve fertility rates (17,49). Aromatase inhibitors, absent under normal circumstances, are found to be present in ectopic endometrial tissue; this may have a direct impact on oestradiol levels and implantation rates in endometriosis patients (50,51). Current treatment of endometriosis-associated infertility focuses on improving fecundity by removing or reducing ectopic endometrial implants, thus restoring normal pelvic anatomy (47). There are several possibilities that practitioners can adopt when facing this kind of events, depending on the particular case and the patient: expectant management, medical treatment and/or surgical treatment (48,52). The reality is that the optimal method of choice to treat endometriosis-associated infertility is an individualized decision that should be made on the basis of the specificity of the patient (18,50). CONCLUSIONS Despite lower ovarian response, reduced embryo quality and impaired implantation in moderate/severe cases, endometriosis patients have been able to obtain IVF/ICSI success rates similar to those with tubal factor-related infertility. As it has been widely explained, the combination of aggressive but controlled ovarian hyperstimulation, appropriate hypophysis suppression and efficient surgery before initiating cycles seemed to be crucial and significantly efficient for IVF/ICSI success on patients suffering from endometriosis (50,52). Within the last few years, scientific knowledge have made it possible to develop certain clinical improvements that have surely opened new possibilities for endometriosis patients. REFERENCES 1. Available from: https://www.womensmd.org/wp-content/uploads/2017/06/Endometriosis-causing-heavy-periods-with-blood-clots.jpg [Cited November 4 2017].
2. Kennedy S, Bergqvist A, Chapron C, d’Hooghe T, Dunselman G, Greb R, et al. ESHRE guideline for the diagnosis and treatment of endometriosis. Hum Reprod. 2005;20:2698–704. 3. Johnson NP, Hummelshoj L, World Endometriosis Society Montpellier Consortium. Consensus on current management of endometriosis. Hum Reprod. 2013;28:1552–68. 4. Vercellini P, Viganò P, Somigliana E, Fedele L. Endometriosis: pathogenesis and treatment. Nat Rev Endocrinol. 2014;10(5):261–75. 5. Giudice L. Endometriosis. N Engl J Med. 2010;362:2389–98. 6. Practice Committee of the American Society for Reproductive Medicine. Endometriosis and Infertility. Fertil Steril. 2006;14:S156–60. 7. Bulletti C, Coccia ME, Battistoni S, Borini A. Endometriosis and infertility. J Assist Reprod Genet. 2010; 27:441–7. 8. Koninckx P, Ussia A, Adamyan L, Wattiez A, Donnez J. Deep endometriosis: definition, diagnosis, and treatment. Fertil Steril. 2012;98(3):564-571. 9. Burney R, Giudice L. Pathogenesis and pathophysiology of endometriosis. Fertil Steril. 2012;98(3):511-519. 10. Lebovic D, Mueller M, Taylor R. Immunobiology of endometriosis. Fertil Steril. 2001;75(1):1-10. 11. Sourial S, Tempest N, Hapangama D. Theories on the Pathogenesis of Endometriosis. Int J Reprod Med. 2014; 179515.1-9. 12. Nyholt DR, Low SK, Anderson CA, Painter JN, Uno S, Morris AP et al. Genome-wide association meta-analysis identifies new endometriosis risk loci. Nat Genet. 2012;44(12):1355–1359. 13. Pagliardini L, Gentilini D, Vigano’ P, Panina-Bordignon P, Busacca M, Candiani M et al. An Italian association study and meta-analysis with previous GWAS confirm WNT4, CDKN2BAS and FN1 as the first identified susceptibility loci for endometriosis. J Med Genet. 2013;50:43–46. 14. Aghajanova L, Hamilton A, Kwintkiewicz J, Vo KC, Giudice LC. Steroidogenic enzyme and key decidualization marker dysregulation in endometrial stromal cells from women with versus without endometriosis. Biol Reprod. 2009;80:105 –114. 15. Wei Q, St Clair JB, Fu T, Stratton P, Nieman LK. Reduced expression of biomarkers associated with the implantation window in women with endometriosis. Fertil Steril. 2009;91: 1686–91. 16. Practice Committee of the American Society for Reproductive Medicine. Endometriosis and infertility: a committee opinion. Fertil Steril. 2012; 98: 591-598. 17. Surrey ES. Endometriosis and assisted reproductive technologies: maximizing outcomes. Semin Reprod Med. 2013;31:154–63. 18. Barnhart K, Dunsmoor-Su R, Coutifaris C. Effect of endometriosis on in vitro fertilization. Fertil Steril. 2002;77:1148 –1155. 19. Kuivasaari P, Hippelainen M, Anttila M, Heinonen S. Effect of endometriosis on IVF/ICSI outcome: stage III/IV endometriosis worsens cumulative pregnancy and live-born rates. Hum Reprod. 2005;20:3130–3135. 20. Opøien HK, Fedorcsak P, Omland AK, Abyholm T, Bjercke S, Ertzeid G, et al. In vitro fertilization is a successful treatment in endometriosis-associated infertility. Fertil Steril. 2012;97:912–8. 21. Karita M, Yamashita Y, Hayashi A, Yoshida Y, Hayashi M, Yamamoto H et al. Does advanced-stage endometriosis affect the gene expression of estrogen and progesterone receptors in granulosa cells? Fertil Steril. 2011;95:889 – 894. 22. Harb H, Gallos I, Chu J, Harb M, Coomarasamy A. The effect of endometriosis on in vitro fertilisation outcome: a systematic review and meta-analysis. BJOG. 2013;120:1308–20. 23. Revelli A, Pettinau G, Basso G, Carosso A, Ferrero A, Dallan C et al. Controlled Ovarian Stimulation with recombinant-FSH plus recombinant-LH vs. human Menopausal Gonadotropin based on the number of retrieved oocytes: results from a routine clinical practice in a real-life population. Reprod Biol Endocrinol. 2015;13:77. 24. Tan CW, Lee IH, Choolani M, Tan HH, Griffith L, Chan J. Endometriosis, endometrium, implantation and fallopian tube. Hum Reprod. 2013;28:206-226. 25. Somigliana E, Infantino M, Benedetti F, Arnoldi M, Calanna G, Ragni G. The presence of ovarian endometriomas is associated with a reduced responsiveness to gonadotropins. Fertil Steril. 2006;86:192–196. 26. Benaglia L, Bermejo A, Somiglianaetal E. Invitrofertilization outcome in women with unoperated bilateral endometriomas. Fertil Steril. 2013;99:1714–1719. 27. Suzuki T, Izumi SI, Matsubayashi H, Awaji H, Yoshikata K, Makino T. Impact of ovarian endometrioma on oocytes and pregnancy outcome in in vitro fertilization. Fertil Steril. 2005;83:908–913. 28. Almog B, Shehata F, Sheizaf B, Tan SL, Tulandi T. Effects of ovarian endometrioma on the number of oocytes retrieved for in vitro fertilization. Fertil Steril. 2011;95:525–527. 29. Hooghe MD, Denys B, Spiessens C, Meuleman C, Debrock S. Is the endometriosis recurrence rate increased after ovarian hyperstimulation? Fertil Steril. 2006;86:283–290. 30. Benaglia L, Somigliana E, Santi G, Scarduelli C, Ragni G, Fedele L. IVF and endometriosis-related symptom progression: insights from a prospective study. Hum Reprod. 2011;26:2368–2372. 31. Mehedintu C, Plotogea M, Ionescu S, Antonovici M. Endometriosis still a challenge. J Med Life. 2014;7:349-57. 32. Surrey ES, Silverberg KM, Surrey MW, Schoolcraft WB. Effect of prolonged gonadotropin-releasing hormone agonist therapy on the outcome of in vitro fertilization-embryo transfer in patients with endometriosis. Fertil Steril. 2002;78:699–704. 33. Lessey BA. Medical management of endometriosis and infertility. Fertil Steril. 2000;73:1089–1096. 34. Tei C, Maruyama T, Kuji N, Miyazaki T, Mikami M, Yoshimura Y. Reduced expression of alphavbeta3 integrin in the endometrium of unexplained infertility patients with recurrent IVF-ET failures: improvement by danazol treatment. J Assist Reprod Genet. 2003;20:13-20. 35. Ziegler D, Gayet V, Aubriotetal FX. Useoforalcontraceptives in women with endometriosis before assisted reproduction treatment improves outcomes. Fertil Steril. 2010;94:2796–2799. 36. Furness S, Yap C, Farquhar C, Cheong Y. Pre and post-operative medical therapy for endometriosis surgery. Cochrane Database Syst Rev. 2004:CD003678. 37. Opøien HK, Fedorcsak P, Byholm T, Tanbo T. Complete surgical removal of minimal and mild endometriosis improves outcome of subsequent IVF/ICSI treatment. Reprod Biomed Online. 2011;23:389 – 395 38. Barri PN, Coroleu B, Tur R, Barri-Soldevila PN, Rodríguez I. Endometriosis-associated infertility: surgery and IVF, a comprehensive therapeutic approach. Reprod Biomed Online. 2010;21:179–185. 39. Bianchi PH, Pereira RM, Zanatta A, Alegretti JR, Motta EL, Serafini PC. Extensive excision of deep infiltrative endometriosis before in vitro fertilization significantly improves pregnancy rates. J Minim Invasive Gynecol. 2009;16:174–180. 40. Surrey ES, Schoolcraft WB. Does surgical management of endometriosis within 6 months of an in vitro fertilization–embryo transfer cycle improve outcome? J Assist Reprod Genet. 2003;20:365–70. 41. Khamsi F, Yavas Y, Lacanna IC, Roberge S, Endman M, Wong JC. Exposure of human oocytes to endometrioma fluid does not alter fertilization or early embryo development. J Assist Reprod Genet. 2001;18:106-109. 42. Benschop L, Farquhar C, Van der Poel N, Heineman MJ. Interventions for women with endometrioma prior to assisted reproductive technology. Cochrane Database Syst Rev. 2010;11:CD008571. 43. Elter K, Oral E. Surgical treatment before assisted reproductive technologies. Semin Reprod Med. 2014; 32(04): 253-261. 44. Barcelos ID, Vieira RC, Ferreira EM, Martins WP, Ferriani RA, Navarro PA. Comparative analysis of the spindle and chromosome configurations of in vitro-matured oocytes from patients with endometriosis and from control subjects: a pilot study. Fertil Steril. 2009:92;1749–1752. 45. Salim R, Riris S, Saab W, Abramov B, Khadum I, Serhal P. Adenomyosis reduces pregnancy rates in infertile women undergoing IVF. Reprod BioMed Online. 2012;25:273– 277. 46. Dmowski WP, Pry M, Ding J, Rana N. Cycle specific and cumulative fecundity in patients with endometriosis who are undergoing controlled ovarian hyperstimulation-intrauterine insemination or in vitro fertilization-embryo transfer. Fertil Steril. 2002;78:750-756. 47. Macer ML, Taylor HS. Endometriosis and infertility: a review of the pathogenesis and treatment of endometriosis-associated infertility. Obstet Gynecol Clin North Am. 2012;39: 535-549. 48. Ruiz Flores FJ, Garcia-Velasco JA. Is there a benefit for surgery in endometrioma associated infertility?. Curr Opin Obstet Gynecol. 2012;24(3):13640. 49. Hadman M, Omar SZ, Dunselman G, Chenog Y. Influence of endometriosis on assisted reproductive technology outcomes: a systematic review and meta-analysis. Obstet Gynecol. 2015;125(1):79-88. 50. Wahd SA, Alaf SK, Al-Shawaf T, AlTawil NG. Ovarian reserve markers and assisted reproductive technique (ART) outcomes in women with advanced endometriosis. Reprod Biol Endocrinol. 2014;12:120. 51. Kawwass JF, Crawford S, Session DR, Kissin DM, Jamieson DJ. Endometriosis and Assisted Reproductive Technology: United States Trends and Outcomes: 2000–2011. National ART Surveillance System Group. Fertil Steril. 2015;103(6):1537-43. 52. D Xiyuan, L Xiuhua, Wang R, Zhang H. The impact of endometriosis on IVF/ICSI outcomes. Int J Clin Exp Pathol. 2013;6(9):1911-1918. Authors: Javier del Río, Noemi Díaz and Belén Gómez  Figure 1. Blastocyst. Modified from https://i.ytimg.com/vi/dvTXEGcNFZg/sddefault.jpg INTRODUCTION What is cryopreservation? The first successful in vitro fertilization (IVF) treatment was in 1978. Since that, there have been a remarkable number of advances in assisted reproductive technologies (ART). Initially, all available embryos were transferred in IVF treatments owing to its low success rate. However, improvements on clinical and laboratory aspects led not only to increased pregnancy rates, but also to increased risk of multiple pregnancies. To prevent this, fewer embryos are transferred and leftover embryos are cryopreserved for potential future cycles use (1). The first pregnancy resulting from transferring a thawed cryopreserved human embryo was reported in 1983 in Australia (2), and the first live birth following embryo cryopreservation was reported in 1984 in The Netherlands (3). Subsequently, the need for an effective cryopreservation program arose from rapid development and improvements of assisted reproductive technology protocols (1). Cryopreservation is a method that requires cells and embryos to be exposed to non-physiological ultra-low temperatures (from -20°C to -196°C) (Fig.2). It aims to achieve “cryogenic suspension of life” through multiple steps, although this puts the elements at risk of damage or “cryoinjury” during temperature changes and phase transitions. These damages could be chilling injury or ice crystal formation, for instance, as a result of the water exchange between the intra- and extracellular compartments, consequence of dramatic changes in osmotic potential (osmotic shock). Therefore, vitrification requires the use of cryoprotectants to avoid the formation of ice crystals in the cells. Two types of cryoprotectants are necessary: permeating and non-permeating. Mixing both at different relative concentrations reduces intracellular ice formation by removing water from inside the cell. Additionally, it creates an osmotic gradient that helps restrict water movement across the cell membrane, thereby preventing osmotic shock (4). There are two typical methods used for cryopreservation: slow freezing and rapid freezing to achieve vitrification. Vitrification is a term used to describe the transformation of a solution into glass by a dramatic increase in viscosity. This method requires to minimize the time for the sample to be exposed to temperature ranges associated with chilling injury and ice crystal formation. As slow freezing, vitrification causes cell dehydration using cryoprotectants. However, unlike that, there is no attempt to maintain equilibrium on both sides of cell membrane (4). The time frame required to reach ultralow temperatures by vitrification is very brief, almost instantaneous. But, the main concern is the need for using high concentrations of cryoprotectant solutions. These might lead to osmotic shock and it could be toxic to cells, affecting embryo survival. Nevertheless, it is possible to limit toxicity by mixing different cryoprotectants, thereby decreasing their relative concentration and the exposure time of embryos to the solution (5). How efficient is the vitrification? This technique seems to be more attractive than slow freezing because it does not require expensive equipment. It uses small amount of liquid nitrogen and it is a simpler technique to perform once the embryologist has gained enough experience in it (6). A recent research performed by Viladimoiv et al. suggests advantages arising from the freezing and thawing process; the authors hypothesize a theory about “cryo-treatment of the embryo”. According to these authors, as a result of freezing or thawing of the embryos there is a decrease in reactive oxygen species levels, in the rate of mitochondrial DNA mutation and cells detoxification is carried out. Also, the authors describe another mechanism involved in restoring the mitochondrial activity (“jumping effect”) which is part of the physiological process of implantation. However, current available data cannot confirm the hypothesis yet (7).  Figure 2. Cryopreservation of frozen embryos in liquid nitrogen. Advantages and disadvantages of fresh and frozen cycles Nowadays, fresh embryo transfers (ET) are the most common choice in IVF cycles (8). Nevertheless, in the last years, controlled ovarian stimulation has increased the uncertainty on the possible adverse effects of the ovarian hyperstimulation syndrome (OHSS), and also on possible deleterious effects on the endometrium and implications in obstetric and perinatal results (9). In spite of this, recent developments in cryopreservation of oocytes and embryos have led to substantial improvement in IVF outcomes. This resulted in a significant increase in the number of cycles with frozen embryo transfer (FET), which subsequently led to the enhancement of live births rate (10). What are the advantages of a frozen cycle? Ovarian hyperstimulation syndrome The first strong argument for FET strategy is the prevention of OHSS, that results from an increase in vascular permeability (11,8). OHSS is a medical condition affecting the ovaries of some women who take fertility medication to stimulate oocyte growth. OHSS arguably remains a major cause of morbidity in IVF treatment (10). During a fresh cycle, a woman has to undergo hormonal treatment to regulate her menstrual period, to stimulate the development of multiple oocytes (superovulation), and to encourage their maturation (11, 12). However, in a frozen cycle (FC) the patient does not have to go through ovarian stimulation or egg retrieval depending on their circumstances (13). Many people find that FETs are less stressful than fresh cycles because they do not have to worry about oocytes production or whether there will be viable embryos, since those procedures have already been done (9). Deleterious effects on the embryo The optimization of vitrification protocols has reduced the deleterious effects that this process may produce in embryos. Also, it have been observed similar survival and embryo development in FCs compared to fresh cycles (10). Moreover, best quality embryos, morphologywise, can be stored and transferred in a future cycle in better conditions. These data have allowed for an increment of success rates and the confidence of sanitary personnel and patients over FCs (5). Endometrial receptivity The implantation process, one of the crucial steps in the success of ART, requires a reciprocal interaction between the embryo and the endometrium during a small period of time called window of implantation. This interaction involves the embryo, along with its inherent molecular program of cell growth and differentiation, as well as differentiation of endometrial cells into an adequate uterine receptivity (11). Some patients may find easier to turn to FCs, since dealing with the whole process of medication during a normal cycle for ovarian stimulation may result psychologically and emotionally overwhelming. In this regard, FC may also provide a better outcome (3). The importance of an adequate endometrial environment in ART is highlighted in those patients who resort to oocyte donation, where there must be a synchronization between donor and recipient in fresh cycles. Those cases that require an improvement in endometrial receptivity to stimulate implantation of these donor oocytes seem to obtain better results in frozen cycles or in the next fresh cycle (8). Multiplet pregnancies are one of the major safety concerns of IVF due to the increased risk of neonatal and maternal complications. To achieve good results, to would be ideal to select the optimal single embryo to be transferred. Elective single embryo transfer (eSET) is the most effective way to reduce those risky pregnancies (14). How can cryopreservation damage embryos? Upon analyzing some ART studies and results, embryos are able to adapt and develop in a large range of culture media, showing different gene expression models in different environments. Cryopreservation causes stress in embryos and it is known as “hormesis”(5) (Fig.3). However, if the conditions are too unfavorable or toxic, mitochondrial activity is suppressed below the threshold necessary for the development of the embryo, so that implantation in the endometrium will be affected (5).  Figure 3. Mechanism of hormesis (7). Results of embryo transfer in fresh cycles vs. frozen cycles The main current objective of IVF professionals is to improve pregnancy rates in both fresh and frozen-thawed cycles. It is clear that embryo and endometrial receptivity are important factors to promote pregnancy rate. Recently, many researches showed FET can enhance the embryo utilization rate and improve the success rate in contrast to other research lines (15). In Roque et al. systematic meta‐analysis for 633 cycles in women aged 27-33 years old showed that FET resulted in a statistically significant increase in the ongoing pregnancy rate and clinical pregnancy compared with the fresh transfer group (8). Interestingly, the fresh group showed a higher miscarriage rate, but no statistical difference was found when compared with the frozen group. According to these data, it seems that the results of IVF-ICSI cycles can be improved by performing the FET especially in patients with normal or high follicular response. This advantage could be explained thanks to a more physiological preparation of endometrium. Several studies have also shown good results with cryopreservation of all embryos and subsequent FET in those patients most susceptible to develop OHSS (8, 16-19). In contrast, Shavit et al. found lower rates of clinical pregnancy and live births in the vitrified-warmed blastocyst group. The difference in implantation and pregnancy rates could be attributed to a higher proportion of good-quality embryos in the fresh blastocysts transfer group. They suggest that in fresh cycles highest quality blastocyst is selected for transfer and the rest are usually vitrified. Thus, vitrified-warmed blastocysts may have slightly poorer grade after warming and prior to transfer (20). In addition, it is necessary to take into account those cycles with frozen oocytes. Braga et al. found that warmed oocytes transferred in endometrial prepared cycles yield better clinical outcomes than fresh ETs. Indeed, they found that fertilization rate, embryo quality, and developmental competence was decreased in embryos derived from vitrified oocytes (12). Conversely, previous studies have suggested that the results of oocyte vitrification followed by ICSI are not inferior with regard to fertilization, embryo developmental competence, pregnancy rates, and live birth (21, 22, 23). An interesting point found in Braga et al. research is that even with lower embryo developmental quality, warmed oocytes transferred in endometrial prepared cycles resulted in higher pregnancy and implantation rates compared with transfer in fresh cycles. This finding strongly suggests that controlled ovarian stimulation impacts endometrial receptivity, which may be a possible cause of implantation failure after ovarian stimulation (12). Indeed, some studies have suggested that pregnancy rate is inversely related to serum progesterone levels on the day of hCG administration (24-27). It has been demonstrated that elevated progesterone levels on hCG trigger day negatively influence the pregnancy, regardless of the oocyte quality. Raised concentrations of progesterone in the late follicular phase are likely to influence the secretory changes of the endometrium, leading to an asynchrony between embryo and endometrial dialogue, which may result in reduced implantation rate (12). Another issue to consider is the obstetric and perinatal outcomes of frozen-thawed cycles. Maheshwari et al. quantified in a meta-analysis the obstetric and perinatal risks for singleton pregnancies after FET and compared it with those after fresh embryo transfer (28). They indicated better perinatal outcomes in singleton pregnancies after the transfer of frozen‐thawed embryos when compared to fresh IVF embryos. This could be explained by antepartum hemorrhage, very preterm birth (delivery at <32 weeks), preterm delivery (delivery at <37 weeks), small for gestational age, low birth weight (birth weight <2500 g), and perinatal mortality significantly lower in women who received frozen embryos than those transferred with fresh embryos (29, 28). It is important to note that most studies comparing perinatal outcome of singleton births conceived after fresh and cryopreserved ETs included both single and multiple ETs. Therefore, part of the adverse perinatal outcome may be attributed to the vanishing twin phenomenon, which occurs in up to 10% of multiple ETs resulting in a singleton live birth (20). What can we conclude? Elective embryo cryopreservation followed by single FET has attracted increasing attention and has been regarded as a potential innovation of IVF treatment. Choosing the well-selected embryo could further increase the chance of live birth of a eSET, which is of high clinical significance. However, there are great gaps in the literature about the risk/benefit ratio of this strategy, which includes multiple steps of treatment (30). The good outcomes in FC might be associated with having a well‐balanced embryo‐endometrium interaction in FC, and also with lacking controlled ovarian hyperstimulation, which may adversely affect endometrial receptivity during fresh IVF cycles. In addition, when hormone replacement cycles were applied in FETs, estrogen and progesterone were given in physiological doses to mimic natural cycles, while supraphysiological doses of gonadotropins were given in fresh cycles (31). On the other hand, other authors find fresh cycles as the best choice, especially in patients who resort to oocyte donation. In fact, it seems that there is a higher proportion of good-quality embryos in fresh blastocysts compared to vitrified-warmed blastocysts, which may have slightly poorer grade after warming and prior to transfer. (8, 20). In conclusion, each case must be individualized in relation to clinical characteristics of the patients and to oocyte, seminal and embryo quality. By doing so, results will be optimized in each cycle and the chances of achieving a live birth will be highly improved. REFERENCES:
1. Wong KM, Mastenbroek S, Repping S. Cryopreservation of human embryos and its contribution to in vitro fertilization success rates. Fertil Steril. 2014;102(1):19-26. 2. Trounson A, Mohr L. Human pregnancy following cryopreservation, thawing and transfer of an eight-cell embryo. Nature. 1983;305(5936):707-9. 3. Zeilmaker GH, Alberda AT, van Gent I, Rijkmans CM, Drogendijk AC. Two pregnancies following transfer of intact frozen-thawed embryos. Fertil Steril. 1984; 42(2):293-6. 4. Sparks AE. Human embryo cryopreservation-methods, timing, and other considerations for optimizing an embryo cryopreservation program. Semin Reprod Med. 2015;33(2):128-44. 5. Konc J, Kanyó K, Kriston R, Somoskői B, Cseh S. Cryopreservation of embryos and oocytes in human assisted reproduction. Biomed Res Int. 2014;2014:307268. 6. Loutradi KE, Kolibianakis EM, Venetis CA, Papanikolaou EG, Pados G, Bontis I, et al. Cryopreservation of human embryos by vitrification or slow freezing: a systematic review and meta-analysis. Fertil Steril. 2008;90(1):186-93. 7. Vladimirov IK, Tacheva D, Diez A. Theory about the Embryo Cryo-Treatment. Reprod Med Biol. 2017;16:118–125. 8. Roque M, Lattes K, Serra S, Solá I, Geber S, Carreras R, Checa MA. Fresh embryo transfer versus frozen embryo transfer in in vitro fertilization cycles: a systematic review and meta-analysis. Fertil Steril. 2013;99(1):156-62. 9. Gurbuz AS, Gode F, Ozcimen N, Isik AZ.Gonadotrophin-releasing hormone agonist trigger and freeze-all strategy does not prevent severe ovarian hyperstimulation syndrome: a report of three cases. Reprod Biomed Online 2014;29:541-544. 10. Lattes K, Prat M, Robles A, Carreras R, Brassesco M, Checa MA. Ciclos de criopreservación y vitrificación de ovocitos y embriones: indicaciones y transferencia diferida. Guía 21 de Práctica Clínica de la SEF y de la SEGO. 11. Lessey BA. Endometrial receptivity and the window of implantation. Baillieres Best Pract Res Clin Obstet Gynaecol. 2000;14(5):775-88. 12. Braga D, Setti A, Figueira R, Azevedo M, Iaconelli A, Lo Turco E et al. Freeze-all, oocyte vitrification, or fresh embryo transfer? Lessons from an egg-sharing donation program. Fertil Steril. 2016;106(3):615-622. 13. Shapiro BS, Daneshmand ST, Garner FC, Aguirre M, Hudson C. Clinical rationale for cryopreservation of entire embryo cohorts in lieu of fresh transfer. Fertil Steril. 2014;102:3-9. 14. Tobias T, Sharara FI, Franasiak JM, Heiser PW, Pinckney-Clark E. Promoting the use of elective single embryo transfer in clinical practice. Fertil Res Pract. 2016;2(1):1-9. 15. Shen C, Shu D, Zhao X, Gao Y. Comparison of clinical outcomes between fresh embryo transfers and frozen-thawed embryo transfers. Iran J Reprod Med. 2014. Jun;12(6):409–14. 16. Griesinger G, von Otte S, Schroer A, Ludwig AK, Diedrich K, Al-Hasani S, et al. Elective cryopreservation of all pronuclear oocytes after GnRH agonist triggering of final oocyte maturation in patients at risk of developing OHSS: a prospective, observational proof-of-concept study. Hum Reprod. 2007;22(5):1348-1352. 17. D'Angelo A. Ovarian hyperstimulation syndrome prevention strategies: cryopreservation of all embryos. Semin Reprod Med. 2010;28(6):513-518. 18. Griesinger G, Schultz L, Bauer T, Broessner A, Frambach T, Kissler S. Ovarian hyperstimulation síndrome prevention by gonadotropin-releasing hormone agonist triggering of final oocyte maturation in a gonadotropin-releasing hormone antagonist protocol in combination with ‘‘freeze-all’’ strategy: a prospective multicentric study. Fertil Steril. 2011;95(6):2029-2033. 19. Devroey P, Polyzos NP, Blockeel C. An OHSS-Free Clinic by segmentation of IVF treatment. Hum Reprod. 2011;26(10):2593-2597. 20. Shavit T, Oron G, Weon-Young S, Holzer H, Tulandi T. Vitrified-warmed single-embryo transfers may be associated with increased maternal complications compared with fresh single-embryo transfers. Reprod Biomed Online. 2017;35(1):94-102. 21. Trokoudes KM, Pavlides C, Zhang X. Comparison outcome of fresh and vitri- fied donor oocytes in an egg-sharing donation program. Fertil Steril. 2011; 95:1996-2000. 22. Herrero L, Pareja S, Aragones M, Cobo A, Bronet F, Garcia-Velasco JA. Oocyte versus embryo vitrification for delayed embryo transfer: an observational study. Reprod Biomed Online. 2014;29:567-72. 23. Rienzi L, Romano S, Albricci L, Maggiulli R, Capalbo A, Baroni E, et al. Embryo development of fresh ‘versus’ vitrified metaphase II oocytes after ICSI: a prospective randomized sibling-oocyte study. Hum Reprod. 2010;25:66-73. 24. Xu, B., Li, Z., Zhang, H., Jin, L., Li, Y., Ai, J. et al, Serum progesterone level effects on the outcome of in vitro fertilization in patients with different ovarian response: an analysis of more than 10,000 cycles. Fertil Steril. 2012;97 (1321-7.e1-4). 25. Wu, Z., Li, R., Ma, Y., Deng, B., Zhang, X., Meng, Y. et al, Effect of HCG-day serum progesterone and oestradiol concentrations on pregnancy outcomes in GnRH agonist cycles. Reprod Biomed Online. 2012;24:511–520. 26. Bosch, E., Labarta, E., Crespo, J., Simon, C., Remohi, J., Jenkins, J. et al, Circulating progesterone levels and ongoing pregnancy rates in controlled ovarian stimulation cycles for in vitro fertilization: analysis of over 4000 cycles. Hum Reprod. 2010;25:2092–2100. 27. Hamdine, O., Macklon, N.S., Eijkemans, M.J., Laven, J.S., Cohlen, B.J., Verhoeff, A. et al, Elevated early follicular progesterone levels and in vitro fertilization outcomes: a prospective intervention study and meta-analysis. Fertil Steril. 2014;102:448–454.e1. 28. Maheshwari A, Pandey S, Shetty A, Hamilton M, Bhattacharya S. Obstetric and perinatal outcomes in singleton pregnancies resulting from the transfer of frozen thawed versus fresh embryos generated through in vitro fertilization treatment: a systematic review and meta-analysis. Fertil Steril. 2012;98:368–77.e1. 29. Qiao J, Zhang L, Yan L, Zhi X, Yan J. Female Fertility: Is it Safe to "Freeze?". Chin Med J (Engl). 2015;128(3):390. 30. Wei D, Sun Y, Liu J, Liang X, Zhu Y et al. Live birth after fresh versus frozen single blastocyst transfer (Frefro-blastocyst): study protocol for a randomized controlled trial. Trials 2017; 18(253): 1-7. 31. Zhang L, Yan LY, Zhi X, Yan J, Qiao J. Female Fertility: Is it Safe to “Freeze?” Chin Med J. 2015;128 (3):390-7. Authors: Paula Brígido, Roberto de la Fuente and Javier Del Río 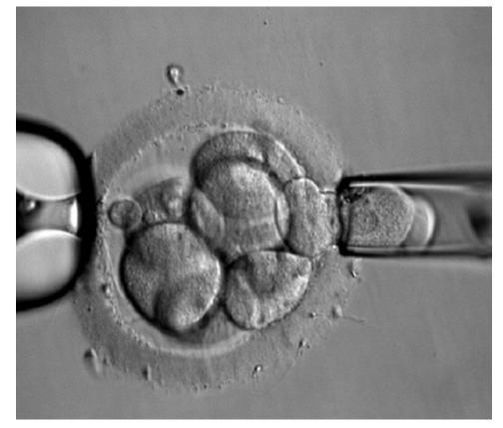 Figure 1. Day 3 embryo biopsy (1) Assisted reproduction technology (ART) can help fertile couples to achieve successful pregnancies. Sometimes, reproductive desires of these couples are affected by the presence of a genetic disease in either partner. In such cases, couples are at a reproductive risk and find themselves in the need of assistance that only ART can provide. Preimplantation genetic diagnosis (PGD) provides an alternative to prenatal diagnosis to detect the specific genetic condition or disease they suffer from, and allows them to avoid passing it on their offspring (2). It requires the analyses of the embryos generated by ART in the in vitro fertilization (IVF) laboratory, by means of accurate and sensitive methodologies such as embryo biopsy, genetics, single cell genomics and, of course, background on prenatal diagnosis and counselling from experts. Clinical application of PGD dates back to the late 60’s, when blastocysts of research animals could be sexed (3) (note that this was already possible ten years before Louis Brown, the first IVF baby, was born in the UK in 1978). At the beginning of the 90’s, early human embryos were sexed before implantation and the first genetic analyses were performed to avoid children inheriting Mendelian diseases. By the end of the century, other nowadays considered basic genetic methodologies were routinely used for preimplantation diagnosis and PGD was applied as a normal procedure to guarantee healthy babies (4). In the present post we aim to give an account of the importance of PGD and the current view of the main clinical approaches for its application. WHEN IS PGD INDICATED? Indications for PGD are multiple and emerge from different motivations. Firstly, the patient may have suffered from a number of terminations due to the embryo having inherited the genetic condition. It could also be motivated by the parents already having a child with a severe genetic disease. In this case they might be willing to avoid passing it on the next one or even looking for a suitable treatment, if possible. However, one of the parents (or both) may be worried about their family history, being aware of the presence of a specific genetic condition, regardless of the type of inheritance. If the parents are carriers of any genetic disease, either an autosomal-dominant disorder like Huntington disease or an autosomal-recessive one like cystic fibrosis, they are at reproductive risk because the resulting embryo may be affected (the probability depending on the specific disorder itself and the way it is inherited) (see [2] for details on inherited conditions). But there are even cases in which motivation is not based on biological but in ethical or religious reasons. Certain families might have serious concerns about going on for abortion of an affected embryo. In such cases, application of PGD may circumvent this kind of ethical conflicts. Applying PGD Broadly speaking, steps for PGD are as follows (2):
PGD vs. PGS Preimplantation genetic screening (PGS) is the general term for a compound of approaches that aim to evaluate the genetic content of the cell, in contrast to genetic tests whose goals are to determine whether an embryo is affected by a specific genetic condition (PGD). Originally termed PGD-AS (preimplantation genetic diagnosis for aneuploidy screening), PGS was developed to confirm the ploidy status of the embryo, searching for possible aneuploidies. Available data suggest that most of miscarriages occurred during the first trimester are a consequence of some sort of aneuploidies (5), and that mainly selected chromosomes were involved in these structural abnormalities (6). Thus, the main approach developed for PGS was the fluorescence in situ hybridization (FISH) for such chromosomes. Types of approaches for PGD in the laboratory Current technical methodologies for preimplantational genetic analyses mainly lie in one of the following:
WHEN TO PERFORM BIOPSY Typical biopsies for PGD (and PGS) are as described as follows:
DAY 3. CLEAVAGE STAGE BIOPSY There is a controversy regarding utility of this type of biopsy. In the cleavage stage biopsy, embryos are biopsied at day 3 when individual cells can be differentiated. This technique entails aspiration of one to two blastomeres to obtain the embryonic genetic material for PGD analysis (13). Following genetic diagnosis, embryo transfer may be performed on blastocyst stage. Embryos are usually selected for biopsy based on morphological criteria. Unfortunately, these do not predict the development potential of the embryo, and so it could fail to progress until blastocyst stage. This would compromise the advantages of using the day-3 approach (14). On the other hand, performing biopsy on the cleavage stage allows embryos to be cultured in vitro until they reach the blastocyst. This means they can be fresh transferred (15), whereas embryos biopsied on day 5 must be vitrified and transferred in a subsequent cycle. How many cells should be removed? The number of cells to be removed in the biopsy is still a controversial issue. Aspirating one cell reduces the cellular mass extracted but it can imply the presence of mosaicism. Conversely, aspirating two cells can reduce the risk of mosaicism, but removing such cellular mass could have consequences on the implantation rate (14). Reported data have shown a dramatic reduction of 39% in the implantation rate in cleavage stage biopsy (16). The authors related it with proportion of the embryo total cellular removed. Whereas around five cells pulled out of the embryo in the trophectoderm biopsy represent 2-3% of the total cell content (expanded blastocyst has 200-220 cells approximately), extraction of a single cell from an eight cell embryo supposes 13% of the total content (16). What do experts say? Cleavage stage biopsy produces different opinions among embryologists because of the presence of mosaicism and the possibility of self-correction of aneuploidies from cleavage to blastocyst stage (17). On the contrary, studies using array-comparative genomic hybridization (array-CGH) technology to analyse genetic abnormalities in day-3 blastomeres and confirming it in trophectoderm biopsy showed concordance between day 3 diagnosis and day 5 reanalysis; Treff and coauthors showed more reliable results for SNP-microarray (96% vs. 83%) and also a lower mosaicism degree (31%) for SNP-microarray samples in a study comparing array technology versus FISH technique (18). These data would support the suggestion of some authors, who proposed that the incidence of mosaicism may have been overestimated in previous studies due to technical inconsistency of the FISH technique (17, 18, 19). At present, this matter remains controversial. Regarding pregnancy rates, in both types of biopsies higher pregnancy rates are obtained comparing with the control group, in which no biopsy was performed (14, 19). To sum up: 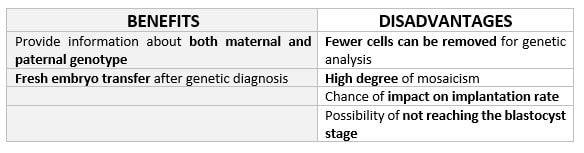 Table 1. Benefits and disadvantages of day 3 biopsy DAY 5. TROPHECTODERM BIOPSY The blastocyst stage is currently supposed to be an optimal time to perform biopsies for PGD/PGS. The combination of improved blastocyst culture, trophectoderm (TE) biopsy, refined cryopreservation techniques, and molecular assays, such as array comparative genomic hybridization that allows for 24-chromosome screening, have led to a renaissance of PGS. TE biopsy will not detect every circumstance in which the embryo is at risk of aneuploidy, but it will detect mosaicism more reliably than cleavage-stage biopsy (which cannot be relied on at all for this purpose) (20, 21). Moreover, when diagnosing monogenic disorders in single blastomere cells using PCR-based protocols, there is a high risk of PCR failure due to either no amplification (allele dropout) or preferential amplification of one of the alleles, potentially resulting in a reduced number of unaffected embryos available for transfer. Increasing the amount of starting DNA template should in principle increase the sensitivity and reliability of genetic diagnosis. Therefore, the biopsy of multiple trophectoderm cells from the blastocyst rather than a single cell from cleavage stage embryos should potentially lead to improved PGD outcome for patients (14). How many cells should be removed? Research to determine the appropriate number of biopsied TE cells in blastocyst biopsies are limited. The exact number of biopsied TE cells is hard to count visually because cells are small and usually remain as a clump. In most studies using comparative genome hybridization or single-nucleotide polymorphism array technology for genetic testing, biopsied TE cells were used for genome amplification and their number was impossible to know. Moreover, some studies showed that removing four to five cells leads to better results. Therefore, the biopsied cell number should be higher in the blastocysts with better TE quality than those with worse characteristics (22, 23). Can biopsies affect blastocyst development and its implantation? Whereas it remains possible that biopsy of cleavage-stage embryos can critically arrest further development through reduction of cell mass, the low miscarriage rates and high term birth rates in the present series, as well as data presently under analysis, suggest that this is not the case for TE biopsy. It can be speculated that the damage to blastocyst development potential caused by TE biopsy would be less for blastocysts with a greater number of TE cells (21, 22). Some experts assured that TE biopsy at the blastocyst stage had no meaningful impact on the developmental competence of the embryo as measured by implantation and delivery rates. This contrasts with the information above-mentioned on the significant reduction in the probability for an embryo to implant and progress up to delivery (16). When combined with TE biopsy and blastocyst vitrification, SNP microarray has resulted in high implantation and low miscarriage rates for some IVF patients (15, 16, 24). Are there any limitations? Owing to the limitations of genetic analysis, most of the biopsied blastocysts need to be cryopreserved by vitrification, and blastocysts with normal results would be transferred in the next frozen cycle. In addition, biopsy of numerous cells from blastocysts with grade B or C may cause damage to the embryo, leading to either its arrest or implantation failure. However, 1-5 cells may be the appropriate biopsied TE cell number to maintain the implantation potential (15, 22). Also, the personnel experience of different embryologists is an influencing factor in this technique. The number of biopsied cells in the blastocyst biopsy is hard to quantify and largely dependent on the experience of embryologist (22). To sum up: 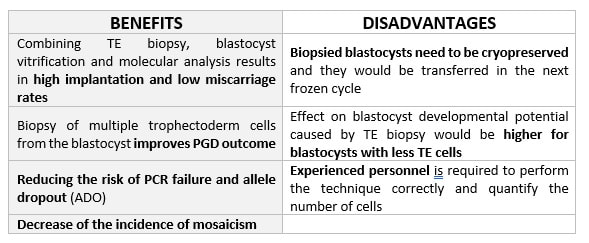 Table 2. Benefits and disadvantages of day 5 biopsy WHAT CAN WE CONCLUDE? The availability of new embryology and molecular techniques allow preimplantation genetic diagnosis laboratories to offer patients at genetic risk the transfer of developmentally competent embryos, unaffected by genetic disease. Cleavage stage biopsy allows for fresh embryo transfer after genetic diagnosis. However, there are reports of high levels of mosaicism when the biopsy is performed on day 3. Trophectoderm biopsy, in turn, provides sufficient material for an effective and more reliable diagnosis in embryos compared to those on cleavage stage. Moreover, it seems that it does not compromise embryo implantation and pregnancy rates in PGD cycles. The drawback for this option is the usual need for cryopreservation and transfer in a different cycle. The offer of PGD in fertility centres has increased over the last decade, primarily due to the progress on the application of diagnostic methods. The choice for either development stage relates to successful outcomes in the clinic, which mainly depend on technical challenges and timing of the developing embryo. For the embryologists, both day-3 and day-5 approaches are supported by evidence, but it will be essential to consider every single aspect of them to evaluate the best option for the laboratory. REFERENCES:
Authors: Shuyana Deba, Isabel Sánchez and Sara Sanz  Figure 1. Benchtop incubators in an IVF laboratory (1) Every procedure carried out in an IVF laboratory, from ovarian puncture and semen capacitation to the embryo transfer, must be performed under specific safety conditions. These standards must be followed to avoid a decrease in gamete/embryo viability (2). From the in vitro culture, gametes and embryos are exposed to diverse artificial situations that do not take place in nature. In vivo, both fertilization and embryo development in the female reproductive tract occur in the complete absence of light. In this environment, other conditions include oxygen (O2) concentration of 2-8%, pH 7.2-7.4, 37⁰C and gradients of diverse nutrients to which the embryo is exposed (3). Changes in temperature, CO2 and O2 pressure, light exposure or volatile organic compounds may adversely affect embryo quality. Also, manipulation of embryos by embryologists is as important as air quality and culture conditions. Each human being is covered by about 1012 bacteria (4), which could contaminate embryo cultures if embryologists do not use the necessary clothing, such as laboratory cap, footwear and mask. Therefore, a daily quality control at different levels should be carried out to obtain good results in IVF cycles. We are going to analyze the effect of some of the elements that can affect germ cells and embryos. LIGHT EFFECT In vivo, mammalian germ cells and embryos are not exposed to light, which might explain why they do not exhibit any protection mechanism against this factor (5,6). In addition, sperm do not have the capacity to repair DNA, unlike oocytes and embryos, which do present some mechanisms for DNA repair (7). Light variables to be considered are intensity, duration and wavelength. It seems clear that photooxidation increases along with light intensity and duration. However, what type of wavelength would be the most harmful for embryos and germ cells? Energy increases when wavelength is shorter (8). Accordingly, artificial cool white fluorescent light has been demonstrated as the most stressful in mouse embryos. Incandescent light, in turn, seems to be less harmful, and the best outcomes are achieved when warm white fluorescent light is used (7). How can light affect the quality of these cells? Indirect effect: Culture and oil photooxidation can affect embryo development (8). In this case, modified components will damage the lipid membranes. Also, if HEPES- or riboflavin-containing media is exposed to light, it results in the formation of hydrogen peroxide, a highly cytotoxic substance (9). Additionally, light can heat up both the plasticware and the oil, resulting in more toxic and damaging components (8,9). Direct effect: Light can potentially compromise the quality of gametes and embryos, by activating stress-related genes or by ionisation, which may also damage the DNA. This phenomenon would cause DNA fragmentation and mutation, as well as an increase in the apoptotic index and change in the number of mitochondria levels (10). How can we avoid this effect? (7) 1) Reducing the exposure time. 2) Using warm white fluorescent light in the lab and green filters on microscopes. 3) Adding antioxidants in the media in order to mitigate damages from ROS. 4) Avoiding riboflavin, which is responsible of the phototoxicity in the media. VOLATILE PARTICLES EFFECT Since the 1990s, IVF laboratory indoor air quality has taken a high relevance. Thence, focused on creating an optimal environment, laboratories have become clean rooms where filtration of particles is performed by using high-efficiency particle arresting (HEPA) filters, and successful chemical air filtration is achieved by removing volatile organic compounds (VOCs) with solid-phase filtration (e.g., potassium permanganate-impregnated, carbon filters) (11). Focusing on VOCs (hydrocarbon-based compounds that are emitted by industries, cleaning products, computers, and microscopes among others), several studies have demonstrated their harmful effect on embryos, initially reported by Boone laboratory on mouse embryo development (11). Moreover, VOCs have been shown to increase DNA fragmentation in human sperm, and they can also have detrimental effects on pregnancy rates (12). Recently, a retrospective study by Munch et al. concluded that, without solid carbon filtration, fertilization, cleavage, and blastocyst conversion rates declined in fresh IVF cycles. Even more, results were found to be even worse in ICSI cycles, probably due to the lack of protective barrier provided by the cumulus cells (13). However, the authors did not observe the same results when embryos had been cryopreserved in an environment with carbon solid filtration but thawed in a laboratory deprived of such systems. The absence of significant changes in cleavage and blastocyst conversion rates, as well as in the proportion of good quality blastocyst developed after thawing suggests that embryos are affected in the peri-fertilization period (13). Also, products like cosmetics emit VOCs, especially perfumes, colognes, and deodorants. They are highly toxic to embryo development in vitro, primarily due to evaporation of their solvent bases (14,15). After analyzing the results of studies determining the toxicity of VOCs, ideal levels should be below 0.2 ppm but preferably zero (12). Personnel must understand the principles of air quality control, including the function of airflows and airlocks, hygiene, dress code and the use of cleaning agents (16). pH EFFECT pH level depends on bicarbonate concentration of culture media and the CO2 concentration of the incubator. However, other factors like altitude and composition of culture media could affect the pH level, too (17,18). Embryos are able to develop over a range of media pH, considering that they possess an intracellular mechanism to regulate its internal level (17,18). However, it is important to control pH variations because they affect development (17). To control pH level outside the incubator some culture media contain buffers like HEPES or MOPS, but long exposure of embryos to these buffers is not recommended (17). Thawed denuded oocytes and embryos are specially sensitive to pH variations because they do not have an inner system to regulate pH (17). So, an increase in the pH of the medium can affect the physiology and development of oocytes and embryos. Thus, acidification of the medium can even affect the fetal weight and size (18). CO2 EFFECT As previously mentioned, CO2 is necessary to control pH level of culture media (17,18). The importance of CO2 was demonstrated in 1985, in a study carried out on hamsters (19). The authors cultured hamster embryos in different CO2 concentrations (5% and 10%). They found a higher rate of blastocysts in those cultured at 10% compared to 5%, which demonstrated differences in embryonic development. This results showed that CO2 level is an important factor for embryo culture (19). The capacity of CO2 to get through cell membranes allows for regulation of the inner pH levels in blastomeres. In other studies, it has been shown that the required CO2 concentration to achieve the optimal pH varies in different species. For instance, the required CO2 level in rats is 7.5%, whereas for humans it is 6.5% (19). O2 EFFECT Some studies have compared different values of O2 concentration in the incubator and they show that a low level (5-6%) improves results when compared to an ambient level (21%). It has been shown that low O2 levels increase implantation, pregnancy and live birth rates (17,20). It seems that a low O2 level reduces ROS in the culture and the presence of volatile particles in the air, although the exact mechanism of action is still unknown (17). TEMPERATURE EFFECT Standard temperature generally used in IVF laboratories is 37⁰C (17,18). However, optimal temperature is unknown because in the female reproductive tract it could be slightly lower, about 36⁰C. On average, temperature of the Fallopian tube is about 1.5⁰C less, whereas the follicular liquid temperature can reach 2-3⁰C lower than core body temperature (17). It is important to control and prevent temperature variations because it can affect meiotic spindle stability and alter embryonic metabolism. It has been shown that an increase of 2⁰C during 20 minutes potentially alters the integrity of the meiotic spindle, which cannot be completely repaired when temperature is set back to 37⁰C. As a consequence of this increase in temperature, embryos express some stress-response genes that compromise development (18). Interestingly, a small decrease in temperature does not have any effect on oocytes, whereas a large difference can be severely harmful for the meiotic spindle (18). CULTURE MEDIA EFFECT Nowadays, there exist two kinds of culture media: one-step media and sequential media (with different compositions for days 0-3 and 3-6) (17,21). All culture media are similar in composition; they contain energy substrates like glucose, pyruvate or lactate, and both organic and inorganic salts, which must be balanced accordingly. Culture media also contain amino acids in different proportions. The exact composition of amino acids in culture media is unknown. One of the most important problems related to the presence of amino acids is the ammonium generated as a product of metabolism. Ammonium has negative effects on embryo and fetal development. To avoid this problem, some culture media contain glutamine, which reduces ammonium production (17,21,22). Also, culture media can be supplemented with macromolecules and other components like HSA, α and β globulins, growth factors, vitamins, lipids, nucleotides, cytokines and hormones (17,22). What can we conclude? There are many parameters that should be kept in mind in order to maintain the optimal conditions for both gamete and embryo development in an IVF laboratory. In vitro, cells and embryos are exposed to different stress situations that must be minimized. Therefore, a routine control at different levels needs to be performed, so that the environment in the laboratory is adapted to resemble the reproductive tract and the intrauterine conditions. REFERENCES:
Authors: Paula Brígido, Shuyana Deba, Javier Del Río and Isabel Sánchez  Figure 1. Assisted hatching performed on an embryo (1) Implantation is one of the key steps in human reproduction, and hatching of the blastocyst is a critical point in the sequence of physiological events that lead to implantation (2). It has been estimated that only 15-20% of embryo transfers culminate in implantation, and as a consequence clinical pregnancy and live birth rates are quite low (3, 4). The potential of an embryo to implant is related to its own nature, the quality of gametes and the endometrial receptivity. But failure in blastocyst hatching is also an important factor that prevents implantation (2, 3, 4). With the aim of solving this problem and in order to improve both implantation and pregnancy rates after Assisted Reproductive Technology (ART) procedures, scientists developed assisted hatching (AH) (2, 3, 4). AH consists of an artificial alteration of the zona pellucida (ZP) either by slimming or breakage. This technique was first described on 1988 by Cohen et al., who reported the first pregnancy after AH (4), and successive works have since shown its efficiency (2, 3). Because AH does not seem to present clear advantages to all patients, it should not be applied, in principle, to all of them as a rule (3). Some studies have shown that AH has effectively improved ART outcomes in patients with bad prognosis, like those with a history of 2+ implantation failures, bad embryo quality, aged patients (38+), patient with frozen embryos or those who have oocytes with a thick ZP (2, 3, 4, 6). ART have some actual consequences on gametes and embryos, one of them being hardening of the ZP by the use of certain culture media or by cryopreservation. These and other negative effects may hamper blastocyst hatching, which might be solved through AH. Overall, this approach might be useful considering the early implantation window in women treated with exogenous gonadotrophin stimulation, compared to a natural cycle. Additionally, benefits from AH include the possibility of ZP breakage improving the embryo-endometrium communication (5). In order to help the embryo hatch through the ZP before implantation, a variety of techniques over the years have been developed (7). As a commonality for all of them, it is important to minimize the time of the embryo being outside the incubator and to optimize the methodologies to reduce both pH and temperature variations (3). MECHANICAL ASSISTED HATCHING (PARTIAL ZONA DISSECTION) This method does not require any new equipment, and it can be easily performed by an embryologist handling a micromanipulator (8). During this procedure, the embryo is held by the suction exerted by the holding pipette, while the microneedle is passed through the zona pellucida at the largest visible portion of the perivitelline space and introduced in a tangential trajectory to the embryo. Then, in order to tear the ZP apart, the embryo is released from the holding pipette and rubbed against it (7). However, one of the main disadvantages of this technique is the difficulty of creating a hole of significant size without abruptly damaging the embryo. Moreover, it is not a fast procedure, which means embryos need to remain outside the incubator for a long time, and so this may affect development of the transferred embryos (7, 9). ACID TYRODE'S ASSISTED HATCHING For this procedure, first the embryo is stabilized with the holding pipette. Second, a pipette containing acid Tyrode is approached to the embryo, oriented at the 3 o`clock position, next to an area of empty perivitelline space. Finally, the acidic solution is gently expelled over a small area (~30 µm Ø) and immediately washed away (7). This technique allows the formation of a hole of significant size; nevertheless, the exposure to acid is problematic, since this compound may be potentially embryotoxic. Furthermore, this technique requires practice and expertise from the operator (7, 10). LASER-ASSISTED HATCHING Laser-assisted hatching (LAH) is designed for easy positioning of the embryos, focus and shooting. It can be executed with a single click of the mouse controller. The diameter of the drilled holes vary between 5 and 10 µm according to how many shots are performed and the irradiation time (7). From the technical point of view, this method is easier, better controlled and more precise. Without physical contact with other embryos, the procedure can be completed faster than other methods. So, time of the embryos out of the incubator is shorter than for other techniques (9). Even though the equipment may result expensive, the laser-assisted technique seems to represent the lowest potential risk for the embryo, and it is relatively simple to perform with consistency between operators (3). EFFICACY OF AH Although it has been performed for more than 20 years, up to date results are still inconclusive. Ma and co-authors performed one of the first trials to determine the overall effect of this technique. They concluded that implantation rates could be enhanced by performing ICSI along with AH, but differences were not statistically significant (11). Two recent meta-analyses evaluating potential benefits of this technique have reported significant heterogeneity among results (12, 13), suggesting that effects of AH may differ depending on specific patient features (14). Most researchers support the hypothesis that this technique improves clinical pregnancy rates in patients with previous failed IVF cycles or poor prognosis. However, there is insufficient evidence to affirm that AH improves live-birth rates in these populations, and so it remains uncertain whether AH is beneficial to other patients (6, 14, 16, 17). IS THERE AN INCREASE IN MULTIPLE GESTATION RATE? Even though certain reports associate artificial manipulation of the zona pellucida with multiple pregnancies, there is actually insufficient evidence to support an increased risk of monozygotic twinning after AH. In fact, the overall rate of monozygotic twin pregnancy in IVF with AH is less than 1% (6, 16). MISCARRIAGE AND CONGENITAL MALFORMATIONS RATES It has been reported that this technique may enhance implantation of abnormal embryos. Thus, the lower live birth rate observed, which is related to the high number of pregnancies ending in early miscarriage, may be due to chromosomal abnormalities of the embryos (17). However, Ma et al. performed cytogenetic tests on miscarriaged embryos and umbilical cord blood from newborn infants, where they found a similar incidence of major congenital malformations in ICSI-born patients compared to the general population (11), thus ruling out any direct relationship of AH and miscarriage. EFFECTS OF ASSISTED HATCHING ON FROZEN CYCLES Data show that cryopreservation may induce zona hardening as well as advanced female age and in vitro culture conditions (18). The answer to this problem could be AH, but results obtained on implantation and pregnancy rates after AH for frozen embryo transfer cycles are controversial, as well as for fresh IVF cycles (19). These discrepancies may be attributed to the type of AH, the extent of ZP microdissection, the number of patients and criteria for their selection, or even the quality and stage of embryos selected for AH performance. In an earlier study, Primi et al. (2004) were unable to show any specific advantage of LAH in cryopreserved embryos. In this study, no embryo selection was reported, so this could explain why the implantation rates observed were lower than those achieved when embryos were selected (20). Ng et al. (2005) also found a negative effect of LAH on frozen-thawed embryos in their randomised study (implantation rates: 9.0% vs. 12.5%; pregnancy rates: 6.8% vs. 15%). Although a subgroup analysis showed a higher implantation rate, differences were not statistically significant when LAH was performed on embryos with zona thickness of 1.6 mm (21). In contrast, other studies using similar techniques of LAH were able to show improvement in implantation and pregnancy rates in the LAH group. Such was the case in a study by Balaban and co-authors (2006), in which their data revealed that implantation rates (20.1% vs. 9.9%) and pregnancy rates (40.9% vs. 27.3%) were significantly higher in the group in which embryos had been subjected to LAH before transfer, as compared to those from the control group (22). Valojerdi et al. (2008) in turn showed that LAH increased significantly both implantation and pregnancy rates in embryo cryopreservation cycles (23). These conclusions resemble those by Kanyo and co-authors (2016), who compared clinical pregnancy rates after using LAH technique on day-3 frozen-thawed embryos, and found a higher pregnancy rate after applying the LAH procedure (24). CONCLUSIONS Reports on patient populations are found that show benefits from AH, which include patients whose embryos present a thick zona, with elevated FSH, over 38 years old and cryopreserved cycles. It seems that the laser-based method is the most used among professionals and the most effective, although differences between are found between studies. However, no significant data show any increase in clinical pregnancy rates that could be translated into an encouragement of this technique to be routinely performed. It should also be noted that some studies do not include specific important confounding factors, such as patient socioeconomic status, parity, duration of infertility, number of repeated failed cycles, embryo quality and/or smoking and alcohol intake, which might cause residual variance in the obtained results. Additional limitations of the study include the lack of information on the type of assisted hatching (mechanical, chemical, or laser), which may have changed over time or between clinics. To learn more about this topic, visit our ‘VIDEOS’ section, where you will find videos about mechanical, chemical and laser hatching. REFERENCES
Authors: Shuyana Deba, Javier Del Río, Isabel Sánchez and Sara Sanz 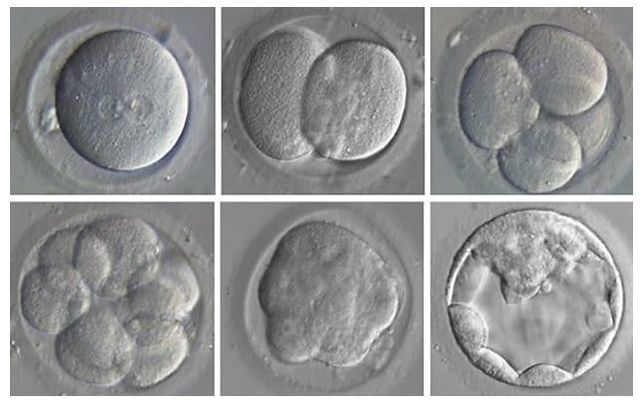 Figure 1. Embryonic development (1) Fertilization is a sequence of coordinated events that results in the metabolic activation of the ootid (nearly mature oocyte) and triggers cleavage of the zygote (2). Nowadays, in assisted reproduction laboratories cleavage can be evaluated in vitro and in real time. Once in vitro fertilization (IVF) has been accomplished, early development of the embryo can be recorded by using time-lapse systems (TLP) (3). This approach makes it possible to evaluate morphology, including dynamic parameters, based on the uninterrupted culture of the embryo, which also allows for a better embryo selection, thus rising pregnancy rates (4). Even so, there are still clinics all over the world that select embryos for transfer using light microscopy, which means the use of a conventional incubation method (5). CRITERIA FOLLOWED FOR EMBRYO CLASSIFICATION It is known that an international consensus is needed in the way embryos are assessed and described. The following standardized criteria is from Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology, 2011 and includes ‘minimum standards’ for oocyte and embryo morphology scoring (6): the current expected observation for embryo development is 4 cells on day 2 and 8 cells on day 3 after fertilization (day 0). Moreover, embryos with <10% fragmentation, stage-specific cell size and not multinucleated are considered of good quality (6). According to this consensus, scoring for day 4 (morula stage) regards as good embryos those that enter into a fourth round of cleavage, which implies evidences of compaction that virtually involve the whole volume of the embryo (6). Finally, on day 5 blastocysts are to be observed expanded with: a prominent inner cell mass (ICM) consisting of many cells, compacted and tightly adhered together; and a trophectoderm (TE), forming a cohesive epithelium (6). Nevertheless, these parameters do not restrict laboratories to annotate further observations in order to select the best embryo for transfer (6). BEST DAY TO PERFORM EMBRYO TRANSFER One of the most important aspects that influence the success of ART is embryo transfer from the culture medium to the uterus (7). This has been a controversial subject that still generates quite some doubts. Morphological evaluation of embryos is sometimes a subjective process, and it provides limited information on the possible genetic abnormalities that embryos may have (8). Currently, there exists a great controversy on the optimal moment to carry out embryo transfer.
IN WHICH CASE DOES THIS TRANSFER USUALLY TAKE PLACE? Day 2 transfer is usually indicated in cases of poorly responding patients. Indeed, it is also indicated when the sperm, oocyte and/or embryos are also of low quality and/or number (9, 10, 11). WHAT DO EXPERTS SAY? Several retrospective studies have compared embryo transfer on day 2, day 3 and day 5 after oocyte recovery, all of which presented conflicting results. A study performed by Mahdavi et al. among poor responder patients revealed no clinical differences between day-2 and -3 embryo transfer (10). However, this study found that pregnancy rates per oocyte retrieval and embryo transfer were significantly higher in the day-2 embryo transfer group compared to day 3 group. It is worth mentioning that other investigators did not find significant differences in pregnancy outcomes when they compared embryo transfer on day 2 and day 3 (11, 12). Additional results from other studies have revealed higher clinical and ongoing pregnancy rates after embryo transfer on day 2 than on day 3 in poor responders. This suggests that the occurrence of miscarriage can be reduced by restricting embryo culture to only 2 days, which could also provide an alternative for managing poorly responding patients (11). That is the reason why embryo transfer on day 2 is still performed in many IVF centres; there is an actual risk of compromising the viability of embryos by prolonged in vitro culture in sub-optimal conditions, with an increased risk of obtaining no blastocysts to replace on day 5 (9, 13, 14). Even though there seems to be a large number of benefits for these patients, certain disadvantages that may potentially occur must also be taken into account, as it can be seen below (Table 1). 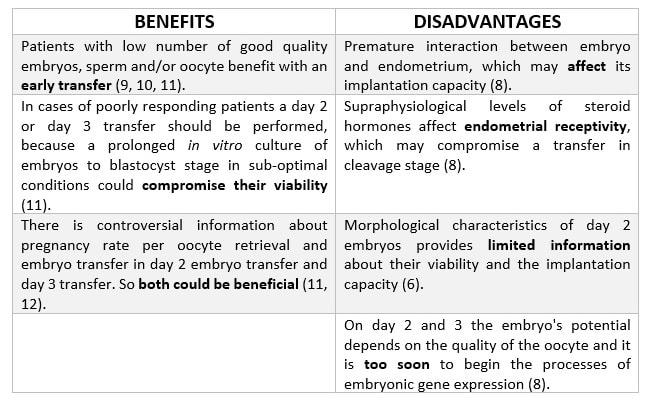 Table 1. Benefits and disadvantages of transferring D+2
PATIENTS IN WHICH TRANSFER D+3 SHOULD BE PERFORMED There exists no criterion to select patients who should be transferred on D+3. Traditionally, embryo transfer has been performed on cleavage stage, so the chosen day was D+3 of embryo development (7). Generally speaking, embryo transfer was carried out on this day in all patients, until a culture medium was developed that allowed to keep embryos in the laboratory for 5-6 days, and with the exception of the cases previously mentioned (11). SCIENTIFIC LITERATURE TO SUPPORT D+3 AS THE BEST DAY FOR EMBRYO TRANSFER Many studies show contradictory results on whether it is better to transfer on D+2 or D+3. However, there are no significant differences as for implantation, clinical pregnancy or live birth rates when comparing transfer on these days. A study by Modares et al. (15) with patients under 40 years old showed a slight improvement in these results when transferring on D+3, although differences were not significant. The authors also showed embryo quality to be worse when the transfer was performed on D+3 than on D+2. Thus, implantation rate has been observed to be higher in D+3, because extending embryo culture for one day allows to discard those embryos that stop their development from D+2 to D+3 (16, 17). Furthermore, it is necessary to consider that there are other external factors that affect embryo development and, consequently, the selection of the best day to transfer. Quinn et al. (18) determined that one of these factors is culture media. Thus, in sub-optimal lab conditions, it would be interesting to transfer on D+2 rather than D+3, in order to spend the shorter time possible in the media. Regarding D+5 transfer, some studies have shown higher implantation rates in embryos transferred on the blastocyst stage compared to those transferred on D+3 (cleavage stage). However, it is necessary to consider that only 25% of embryos reach the blastocyst stage (15); this implies that the number of embryos transferred and vitrified in a cycle is lower than for D+2 and D+3. As a consequence, when considering cumulative pregnancy rates no significant differences are found between transferring on cleavage stage and blastocyst (7). Again, benefits for the patients must be considered along with potential disadvantages (Table 2).  Table 2. Benefits and disadvantages of transferring D+3
It has been observed that transfer on blastocyst stage helps to improve pregnancy rates and reduce the risk of a multiple pregnancy. Why? One reason might be that there is no method to determine whether embryos that initially seem to be of good quality are likely to develop up to blastocyst (19). WHO ARE THE IDEAL PATIENTS? 1. Those with a large number of embryos (20). 2. Those whose day-3 embryos are of good quality (20). 3. Those in which day-1 embryos exhibit pro-nuclei and present a grading profile (20). 4. Young women with good ovarian response (21). 5. Those whose embryos display an early cleavage (22). POTENTIAL BENEFITS OF BLASTOCYST-STAGE TRANSFER vs. CLEAVAGE-STAGE TRANSFER First of all, the new culture media allow us to perform longer incubations in the laboratory, after which the best embryos can be selected with higher accuracy and with lower risk of aneuploidies (23). Moreover, there will exist a better synchronization between the embryo and the mother. Additionally, uterine contractility decreases during the luteal phase (24, 25). The size of these blastocysts is bigger, so some studies have found fewer cases of ectopic pregnancies in comparison to transfers on day 3 (26). A parallel comparison of benefits vs. disadvantages for this procedure can be seen in below (Table 3). 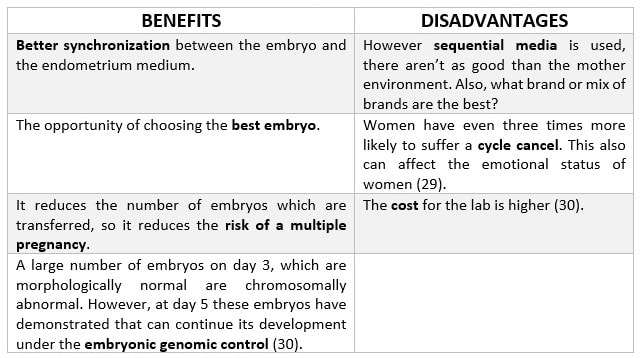 Table 3. Benefits and disadvantages of transferring blastocysts (27) IS IT BETTER TO TRANSFER ON DAY 5 OR ON DAY 6?
The optimal time for embryo transfer depends on a variety of factors, such as the embryo growth speed. However some studies have revealed both implantation and pregnancy rates to be more successful when embryos are transferred on day 5 compared to day 6. This is due to the fact that viability of embryos expanded on day 5 is higher than for those expanded on day 6 (30). In conclusion, it seems difficult to define the most appropriate day for embryo transfer to be applied for each patient. Therefore, every single case must be individually analyzed. In addition, several factors should be taken into account when deciding on the day for embryo transfer, such as maternal age, sperm and oocyte quality or physiological response of the woman and/or the available embryos. By doing so, a good decision can always be made in order to improve implantation and pregnancy rates. REFERENCES
Authors: Shuyana Deba, Javier Del Río and Sara Sanz Special collaboration: Álvaro Martínez Moro 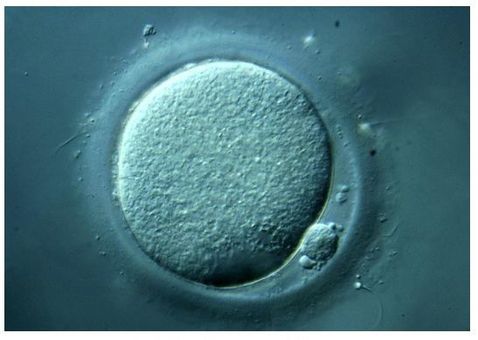 Figure 1. Metaphase II oocyte (1) Infertility affects millions of couples all around the world. In spite of the solutions to their problems reproductive technology can achieve, the efficacy is eventually limited by the number and the quality of the oocytes available from the woman. In actuality, such efficiency is determined by the ovarian reserve, the oocyte quality and the maternal age, among the most important factors (2). Diminished ovarian reserve (DOR) Since ovarian reserve defines the quantity and quality of the primordial follicle pool, diminished ovarian reserve (DOR) indicates a reduction in quantity in women of reproductive age. Consequently, it represents important cause of infertility in many couples. Moreover, DOR may be associated with low pregnancy rates and high pregnancy loss regardless of age, but further research is needed in order to fully understand its implications (3). Advanced maternal age It is well known that women’s fertility declines sharply after age 35 due to several factors, which include specific issues of reproductive organs (uterus and oviducts), general health and decreasing number and quality of oocytes over time. The oocyte pool starts to decline during foetal life and continues within the reproductive life of women. Oocyte quality also decreases as a consequence of the increased rate of aneuploidies observed with age: 74% at the age of 41–42, and up to 93% after the age of 42 (5). Advanced age is too associated with a reduction in the quality of the oocyte cytoplasm (ooplasm), which directly affects oocyte maturation (3). What are the main reasons for this reduction in ooplasm quality? Mitochondria are one of the most important organelles, which are affected in different ways (6,7): - Morphological and functional abnormalities - Mitochondrial swelling - Alterations in mitochondria's cristae - Vacuolization - Alterations of the membrane potential - Alterations of the metabolic pathways in cummulus cells, which may result in impaired mitochondria biogenesis during oogenesis. These effects are due to the higher ratio of mutation consequence of the proximity of these organelles to the respiratory chain, the inefficient repair mechanism and the exposure of histories. How these changes affect oocyte quality (8)? First of all, negative effects on chromosome segregation have been observed as a result of a decreasing ATP concentration (9,10). Additionally, defects have been found in different signalling pathways such as Ca2+ signalling, which affects fertilization and the subsequent embryo development (11). 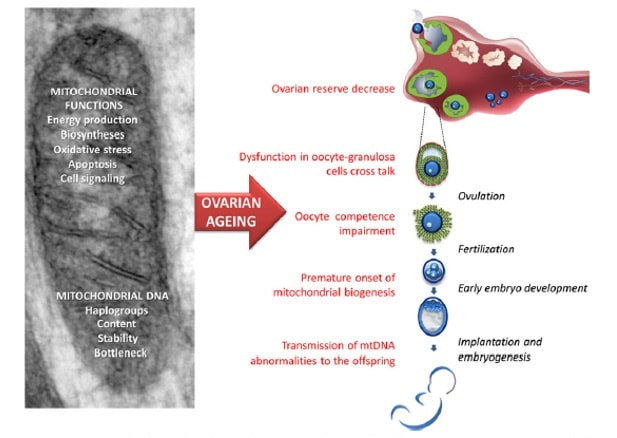 Figure 2. Overview of the multiple implications of mitochondria in ovarian ageing and their deleterious consequences on reproductive function (12) Nevertheless, different mitochondrial haplogroups should be taken into consideration. These have different bioenergetic functions, including production of reactive oxygen species (ROS) and mitochondrial coupling efficiency, aspects that might affect the oocyte longevity (13). Consequently, new techniques are being developed in order to increase the reproductive options in women with oocyte problems. Recently, one of these techniques that have been highly treated in the media is the development of additional viable oocytes from polar body genomes (2). HOW DOES TRANSFER OF POLAR BODY GENOME WORK? Originally, the transfer of polar body has been applied to cases of infertility with a genetic cause, such as the presence of mitochondrial diseases. These cases can be treated with the use of donor oocytes in clinical practice. Additionally, another application is the formation of human metaphase II (MII) oocytes, which increases the number of available oocytes for an assisted reproduction cycle (2). Two specific combined steps are needed. First, the donor oocyte spindle is removed, which requires the utilization of polarized light. Once located, it will be biopsied, obtaining an enucleated oocyte (14,15,16). Secondly, the patient polar body is biopsied, provided elimination of the spindle apparatus has been confirmed. Once both processes have been performed, the last step is the introduction of the polar body genome inside the enucleated oocyte (17). 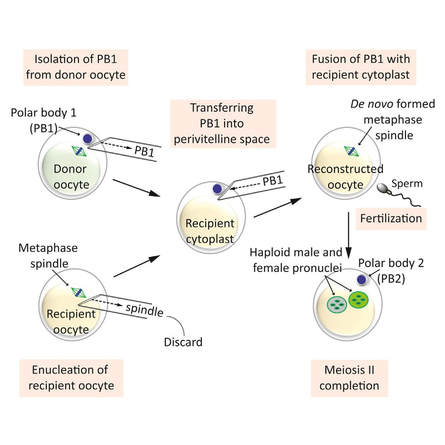 Figure 3. Polar body genome transfer technique (2) FUNCTIONAL HUMAN OOCYTES GENERATED BY TRANSFER OF POLAR BODY GENOMES Hong Ma and his group have tried to test the efficiency and possible limitations of this technique (ref). The main objective to be achieved was the formation of spindles resembling those typical of MII oocytes, including the appropriate chromosome dosage. HOW EFFICIENT IS THIS TECHNIQUE? Although DAPI staining demonstrated that all polar body nuclear transfer (PBNT)-oocytes contained spindle-chromosome complexes, only two of five experimental oocytes formed metaphase spindles similar to intact MII oocytes. This low number may be due to residual meiotic activity in enucleated human MII oocytes, which is sometimes not enough to induce formation of normal MII-like spindles. For a different cohort of oocytes, the rate of successful fertilization was 76%, still slightly lower than control oocytes. Furthermore, 42% of embryos reached blastocyst stage, indicating that most of the PBNT-oocytes were capable of completing the second meiotic division. Short tandem repeat (STR) analysis revealed that two sampled PBNT-blastocysts contained normal diploid chromosomes, determining that these embryos were completely viable. WHAT CAN BE CONCLUDED? • Polar body genome transfer seems to be a significant technique for the improvement of assisted reproductive technology (ART) outcomes and pregnancy rates, particularly for women with decreased ovarian reserve and low response to stimulation. • The cytoplasm from young donor oocytes may reduce incidences of low cytoplasmic oocyte quality. • It could provide an additional technique to support mitochondrial replacement therapy. Nevertheless, this technique is not suitable for women who cannot produce mature oocytes, typical profile of ART patients. Additionally, incidences of aneuploidy resulting from errors in mitosis or in the second meiotic division may still occur because of women advanced age. Larger datasets from this technique are needed to confirm its efficacy and safety. Also, improving preimplantation genetic screening (PGS) is critical before eventual clinical application. REFERENCES:
Authors: Shuyana Deba, Javier Del Río and Sara Sanz  Figure 1: MII oocytes obtained in vitro (1) Some weeks ago, Dr. Hayashi's group published a study in Nature in which they described how to generate functional mouse eggs from cultured stem cells. To better understand this experiment, it is necessary to explain the natural ovarian cycle. Although the experiment was carried out in mice, we believe these experiments could be of great importance in humans in the near future. Therefore, the following description applies to human ovarian cycle. GONADAL DEVELOPMENT IN THE EMBRYO As the yolk sac begins to incorporate into the embryo, germ cells migrate along the dorsal mesentery of the hindgut to the gonadal ridges, which they reach by the end of the fourth or early fifth week of development (2). Around week 7 of gestation, gonadal ridges undergo proliferation to form primitive sex cords, to which primordial germ cells (PGCs) are associated (Fig. 2). In the absence of both expression of the SRY gene product and male sex hormones, differentiation into female organs is determined (3,4). Whereas the origin of the follicular cells of the ovary remains unclear, likely candidates are cells from the coelomic epithelium and the mesonephros. The follicular cells eventually associate with the PGCs to form primordial ovarian follicles (3,4). ORIGIN OF GERM CELLS Human oocytes are derived from round PGCs that can be identified in the wall of the yolk sac as early as 24 days after fertilization. Once the gonad develops into an ovary, PGCs become oogonia, and mitotic divisions go on up to the seventh foetal month, and cease at some point shortly before birth (5). By the 8-9th week after fertilization, some oogonia enter into prophase I of meiosis, thus becoming primary oocytes. Meiosis starts in the deepest part of ovary (medulla) and progresses towards the cortex. Some time after it has been incorporated into a primordial follicle, the oocyte arrests after diplotene, at a particular stage called dictyate or dictyotene. This means the oocyte enters into a long quiescence period that begins before birth and ends up either resuming after the LH surge (leading to ovulation) or in follicular atresia (3). Even though the experiment by Hayashi's group has just been successfully performed in mice, the purpose of the investigation is eventually applying it to humans. 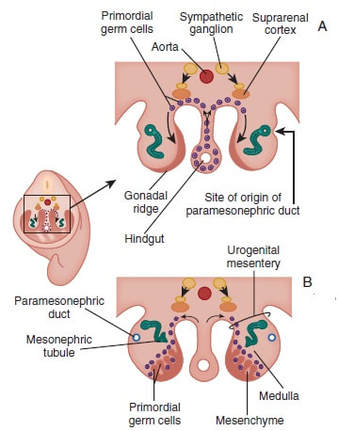 Figure 2: Gonadal development. C: Transverse sections showing the gonadal ridges and the migration of primordial germ cells into the developing gonads; D: transverse section through a 6-week-old embryo, showing the primary sex cords and the developing paramesonephric ducts (3) FOLLICULOGENESIS AFTER BIRTH Once placental oestrogen disappears, gonadotrophin levels begin to increase after birth. This increment will last 12-24 months postpartum in women. Thus, the ovaries remain inactive until the beginning of pubertal stage, moment when they will start showing response to GnRH pulsatility (6). By this time, ovaries contains germ cells at the primordial follicle phase, which are composed of small immature oocytes arrested at dictyate, and surrounded by a single layer of flat squamous granulosa cells (7). WHAT HAPPENS AFTER PUBERTY? Once sexual maturity is reached, multiple primordial follicles are activated from their quiescent state, resuming folliculogenesis by cohorts every menstrual cycle. During follicle maturation, follicles are subjected to activin, AMH, FSH, inhibin, LH, TGFβ and a multitude of other molecules that control the process. At the beginning of this process, as the oocyte resumes meiosis follicular cells proliferate and surround the oocyte, which develops into a primary follicle (7,8,9). Briefly, transition into the secondary or antral follicle stage involves the alignment of the stroma around the basal lamina and the development of an independent blood supply. Follicular cells of the underlying stroma, the granulosa cells, subsequently differentiate into an inner and an outer theca layers, the latter supporting this blood supply. The production of follicular fluid by these cells promotes the formation of the antrum in pre-antral follicles (10). Progression through antral follicle development is promoted by FSH and coincides with (7): - Completion of oocyte growth. - Acquisition of competence to complete meiosis (dependent on fertilization). - Continued granulosa cell proliferation. - Differentiation of cummulus and mural granulosa cells upon formation of the follicular antrum. Response to FSH causes growth of the follicle by proliferation of follicular cells and coalescence of the astral cavities, thus generating a follicle whose volume keeps increasing. The final stage of folliculogenesis takes place when the Graafian follicle reaches its maximum size (around 20 mm diameter). The LH surge triggers the ovulation of a metaphase II-arrested oocyte (along with the cummulus), capable of being fertilized and able to support embryonic development (7). Although there are notable differences between species, in humans, usually only one follicle is capable of completing growth and maturation up to ovulation (preovulatory), due to its more efficient response to FSH in a "competitive manner" against the rest of developing follicles. The one follicle will progress until subsequent fertilization (dominant follicle), whereas the remaining follicles in the cohort will undergo atresia (8). 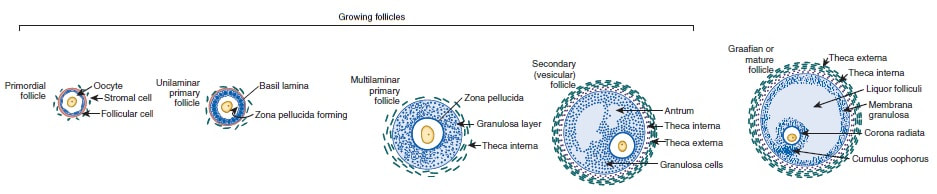 Figure 3. Shematic drawing of development of ovarian follicles (3) ABOUT PLURIPOTENT STEM CELLS For centuries, scientists have investigated how oocytes and sperm develop in mammals. Most of this work has been performed in non-human species, mainly the mouse. Human pluripotent stem cells (PSCs)-derived gamete research presents notable scientific value and potential, both for understanding basic mechanisms of gamete biology and for overcoming clinical problems (11). PSCs are currently used to answer important scientific questions such as the role of specific genes in early germ cell development, involved pathways, interaction between germ cells and supporting somatic cells, or partly or entirely in vitro PSCs-derived eggs and sperm (11). Some of the potential benefits of these new techniques could be (11): - Additional options for assisted human reproduction. - New ways to prevent and treat infertility, genetic diseases and some types of cancer. - Optimization of the number of oocytes obtained from women. "IN VITRO RECONSTITUTION OF THE ENTIRE CYCLE OF THE MOUSE FEMALE GERMLINE" (1) The authors have shown a method to reconstruct the entire process of mouse oogenesis in vitro, using embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) in order to obtain fully functional and mature oocytes. Why does this study mean so much? First of all, this is a big achievement in the field of regenerative medicine, through which it is possible to study the function of certain genes involved in oogenesis. Furthermore, it sets the ground for future similar studies and culture systems in other specie, and as such, it may result in a great advance in reproductive medicine. However, which are the limitations for this approach? The culture system requires somatic cells, which are obtained from embryos. So far, this is an ethical and legal issue in humans. A feasible solution would be to obtain the gonadal somatic cell-like cells from PSCs. The authors found that the differentially expressed genes (DEGs) were down-regulated in in vitro MII oocytes, whereas in vivo MII oocytes showed an increase in the number of transcripts. This maybe a consequence to the fact that, during in vitro growth (IVG) and in vitro maturation (IVM), oocyte development was jeopardized within a subset of (or perhaps all) the oocytes. Why? Misregulation of such genes, related to mitochondrial functions, might attenuate the potential of the in vitro MII oocytes. This problem could be solved by an analysis of the metabolic pathway, which could in turn provide information to refine both IVG and IVM. RESULTS, "PROBLEMS" AND "ACHIEVEMENTS" OF THE EXPERIMENTS (1)  As it can be seen concluded from above, it seems clear that further tests are still needed, which would help measure some aspects of the viability and functionality of this in vitro generated gametes.
REFERENCES 1. Hikabe O, Hamazaki N, Nagamatsu G, Obata Y, Hirao Y, Hamada N et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature. 2016;. 2. Baillet A, Mandon-Pepin B. Mammalian ovary differentiation–A focus on female meiosis. Molecular and Cellular Endocrinology. 2012;356(1-2):13-23. 3. Bolender DL, Kaplan S. Chapter 3 – Basic Embryology. In: Fetal and Neonatal Physiology. 2017. p. 23–39.e2. 4. Parker KL, Schimmer BP. Chapter 8 – Embryology and Genetics of the Mammalian Gonads and Ducts. In: Knobil and Neill’s Physiology of Reproduction. 2006. p. 313–36. 5. Racowsky C, Gelety TJ. Chapter 4 – The Biology of the ovary. Princ Med Biol. 1998;12:77–102. 6. David Vantman B, Margarita Vega B. Fisiología reproductiva y cambios evolutivos con la edad de la mujer. Revista Médica Clínica Las Condes. 2010;21(3):348-362. 7. Hutt K, Albertini D. An oocentric view of folliculogenesis and embryogenesis. Reproductive BioMedicine Online. 2007;14(6):758-764. 8. Green L, Shikanov A. In vitro culture methods of preantral follicles. Theriogenology. 2016;86(1):229-238. 9. Palermo R. Differential actions of FSH and LH during folliculogenesis. Reproductive BioMedicine Online. 2007;15(3):326-337. 10. Gougeon A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Annales d'Endocrinologie. 2010;71(3):132-143. 11. Mathews D, Donovan P, Harris J, Lovell-Badge R, Savulescu J, Faden R. Pluripotent Stem Cell-Derived Gametes: Truth and (Potential) Consequences. Cell Stem Cell. 2009;5(1):11-14. |
Entries
March 2019
Categories
All
 2016-2019. All Rights Reserved by Embryologist Media. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License . |
Embryologist Media

{kind=link}